Riassunto
Il lavoro espone i risultati di uno studio svolto sulla componente macrozoobentonica, eseguito con una metodica di rilievo fotografica su 12 affioramenti rocciosi situati nel Nord Adriatico a diversa distanza dalla costa (1-28 km) e profondità (10-27 m). Le specie individuate dall’esame delle immagini fotografiche (almeno 10 per affioramento) sono state ripartite nei principali grup-pi tassonomici: Porifera, Ascidiacea, Anthozoa e Bryozoa.
Rispetto alla metodica di campionamento distruttiva, eseguita contemporaneamente e sui me-desimi siti, quella fotografica, consentendo ad un livello tassonomico meno definito, ha fornito un quadro complessivo paragonabile, confermando il gradiente costa-largo, la ripartizione degli affioramenti nella cluster analysis e il trend degli indicatori copertura e ricoprimento e, in parte, degli indici di diversità.
Se si soppesano vantaggi e svantaggi, il “metodo fotografico” può rappresentare uno strumen-to essenziale nei programmi di monitoraggio istituzionali in aree protette o di elevato valore ambientale, essendo non distruttivo, consentendo indagini su maggior scala per evidenziare le macroevidenze e fungendo da “early warning” di stress o impatti sull’habitat.
Abstract
Zoobenthic community of North Adriatic rocky outcrops: com-parison between photographic analysis and the destructive Grattage method
Macrofauna assemblages on 12 rocky outcrops in the northern Adriatic Sea located at different distances from the coast (between 1 and 28 km) and depth (between 10 and 27 m) were analyzed by high-resolution digital photography. Macrofaunal species, identified by examining digital pho-tographs (at least 10 each outcrop) were divided into four taxonomic groups: Anthozoa, Bryozoa, Porifera and Ascidicaea. The comparison with direct sampling destructive method, performed on the same sites at the same time, despite the less defined taxonomic level, shows that the photographic method has provided similar results, confirming the gradient between coastal and offshore area, the distribution of outcrops in the cluster analysis and the trend of coverage indicators and – at a reasonable level – diversity indices as well. Considering advantages and disadvantages, the photo-graphic method can be an essential tool, providing a large-scale monitoring planning in institutional monitoring programs, able to highlight habitat degradation in areas of high environmental value.
Introduzione
La costa del Nord Adriatico è un’area fortemente antropizzata, dove l’economia turistica e le attività economiche ad essa legate, quali quelle ricreative in ambito marino costiero, hanno avuto un forte sviluppo nel corso degli ultimi decenni (Adriatic Sea Forum, 2016; CNA, 2015). Tra le attività ricreative, sviluppatesi nel corso degli anni ’90, anche la subacquea ha subito una forte crescita che ha portato molti appassionati a scoprire i fondali prossimi alle città turistiche che si affacciano sulla costa.
Tra i più importanti siti di immersione che si dipanano lungo il tratto costiero, ad una distanza compresa tra poche centinaia di metri e diversi chilometri dalla costa (> 30 km), gli affioramenti rocciosi denominati “tegnùe” rappresentano una delle maggiori attrattive di questo tratto di mare, grazie anche alle profondità relativamente poco elevate, 10-30 m, se si considerano altri siti di immersione mediterranei (Aa.Vv., 2010).
Parallelamente allo sviluppo dell’attività subacquea, sono cresciuti anche gli sforzi economici e scientifici per lo studio delle comunità biologiche di questi biotopi (Mizzan, 1992, 1995; Gabriele et al., 1999; Molin et al., 2003; Casellato & Stefanon, 2008), tanto più dopo l’inserimento di alcuni di questi affioramenti nella rete di siti di importanza comunitaria che formano il network Natura 2000 della Comunità Europea (EX. DIR. 92/43/CEE).
Nell’ultimo decennio sono stati attuati dalla Regione del Veneto e dal Ministero delle Infrastrutture e dei Trasporti – Magistrato alle Acque di Venezia (ora Provve-ditorato alle Opere Pubbliche del Triveneto), tramite il Concessionario Consorzio Venezia Nuova, oltre che dall’ARPAV, studi finalizzati all’approfondimento delle conoscenze sullo stato ecologico di alcuni affioramenti e alla comprensione di quali siano le principali forzanti che guidano i processi di colonizzazione e di crescita delle comunità animali e vegetali (Molin et al., 2010; Curiel & Molin, 2010; Miotti et al., 2014; Curiel et al., 2014; Regione del Veneto-Magistrato alle Acque-Thetis-SELC, 2012; Aa.Vv., 2010).
Gran parte di questi studi sono stati condotti con le tecniche classiche di campio-namento del benthos di substrato duro che si basano sulla raccolta di organismi mediante grattaggio con l’ausilio di una sorbona; tuttavia, molto spesso, nei sub-strati duri è necessario eseguire raschiamenti di aree piuttosto estese per ottenere misure rappresentative, distruggendo una parte rilevante di comunità biologica. Inoltre, la comunità è spesso dominata da organismi raccolti a patches per cui, maggiori sono l’estensione dell’area campionata e il numero di repliche, più rap-presentativo risulterà il campione (Weinberg, 1978; Gambi & Dappiano, 2003). A fronte di queste evidenze e del fatto che gli affioramenti rientrano pienamente nell’habitat di importanza comunitaria 1170 – Scogliere, alcuni di essi, localizzati al largo della città di Chioggia e in prossimità dell’abitato di Caorle, sono protetti da leggi regionali e locali, nonché dalla normativa europea (DIR.92/43/CEE). In generale, ottenere un campionamento rappresentativo dell’habitat di affiora-menti rocciosi del Nord Adriatico può risultare complesso a causa della distri-buzione a patches degli organismi, dell’elevato valore naturalistico delle specie da raccogliere, o per le ridotte dimensioni dell’area da campionare, come nel caso di grotte o piccole secche in mezzo al mare.
Nel corso degli anni ’70-’90, per lo studio degli ambienti batiali, sono state svi-luppate tecniche specifiche di campionamento fotografico e stereo fotografico (Fedra et al., 1979; Christi, 1980; Lampitt et al., 1986; Gage & Tyler, 1991; Pronzato, 1996) e di campionamento mediante videocamera o R.O.V. – Remotely Operated Vehicle.
In seguito allo sviliuppo di queste tecniche di campionamento remoto, alcuni autori sono stati in grado di effettuare, per gli affioramenti anche del Nord Adriatico, stime di abbondanza e di copertura superficiale dai campioni foto-grafici (Molin et al., 2008; Ponti & Mastrototaro, 2006; Ponti et al., 2006; 2011) e hanno provato a correlare questi parametri per ottenere stime di bio-massa come biovolume o peso (Bohnsack,1979; Shears et al., 2003; Molin et al., 2003; 2006a; 2006b; 2009;Molin& Berton, 2007).
Il presente lavoro confronta i risultati di uno studio della comunità macrozoo-bentonica, eseguito con una metodica fotografica, con quelli di un campiona-mento distruttivo, eseguito contemporaneamente sui medesimi affioramenti e transetti (Miotti et al., 2014). Il lavoro fa riferimento ad uno studio più este-so, prommosso dalla Regione del Veneto e dal Ministero delle Infrastrutture e dei Trasporti – Magistrato alle Acque di Venezia tramite il Concessionario Consorzio Venezia Nuova (Studio Op. 499), atto a valorizzare le aree costiere prospicienti le bocche di porto della Laguna di Venezia (Regione del Veneto-Magistrato alle Acque-Thetis-SELC, 2012).
Il presente lavoro rientra nell’ambito dello sviluppo e ricerca di soluzioni non impattanti per il monitoraggio ambientale in mare e, in particolare, dato l’elevato valore ecologico degli affioramenti rocciosi, ha permesso di confrontare dati di campioni prelevati con metodica distruttiva con quelli di campioni fotografi-ci della comunità bentonica.
Area di studio
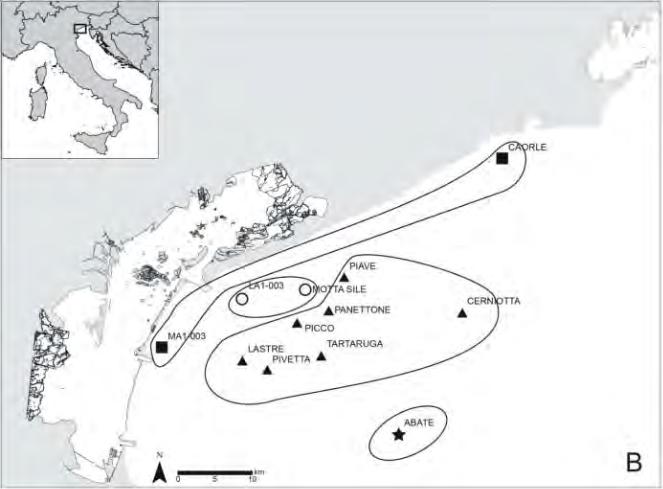
Sono stati indagati 12 affioramenti rocciosi di origine naturale distribuiti al lar-go della costa veneta (Fig. 4) secondo i principali gradienti che caratterizzano le comunità bentoniche, quali la distanza dalle principali fonti terrigene (bocche lagunari e foci fluviali), la profondità, le caratteristiche morfologiche (affiora-menti singoli e compatti, affioramenti costituiti da molti massi, lastrure) e, infi-ne, l’elevazione dal fondale (Tab. 1).
| Tegnùa | Distanza costa (km) | Profondità (m) | Tipologia | Elevazione (m) |
| MA1-003 | 1 | 10 | Rocce sparse | 0,5-1,0 |
| CAORLE | 2,9 | 9-12 | Rocce compatte | 1,0-2,0 |
| LA1-003 | 4 | 13,4 | Rocce sparse | 0,5-2,0 |
| MOTTA SILE | 5,8 | 19 | Rocce compatte | 0,4-2,4 |
| PIAVE | 6,8 | 15-19 | Rocce compatte-lastrure | 3 |
| PICCO | 9,6 | 17-21 | Rocce compatte | 3,0-4,0 |
| PANETTON | 9,8 | 20-21 | Rocce compatte | 2 |
| LASTRE | 10,5 | 19 | Lastrure | 0,5-1,0 |
| PIVETTA | 13,8 | 20 | Rocce compatte-lastrure | 1 |
| TARTARUGA | 15,1 | 19-21 | Rocce compatte | 2 |
| CERNIOTTA | 17,6 | 24 | Rocce compatte | 0,4-2,2 |
| ABATE | 28,8 | 25-27 | Rocce compatte | 2,0-3,0 |
Materiali e metodi
Il disegno sperimentale applicato per lo studio della componente animale è stato di tipo stratificato e ha previsto l’individuazione, attraverso l’analisi dei dati raccolti dalla strumentazione acustica (Multibeam e Side Scan Sonar), di transetti orizzontali lungo la parte superiore degli affioramenti dove eseguire i campionamenti (Aa.Vv., 2010). In ciascun affioramento sono stati eseguiti 50 scatti fotografici random di dimensioni 25 x 20 cm lungo il transetto, per un to-tale complessivo di 600 scatti fotografici; per il campionamento è stata utilizza-ta una macchina fotografica scafandrata con illuminazione artificiale fornita di due lampade laterali e montata su una struttura rigida metallica, per mantenerla sempre parallela e alla stessa distanza dal substrato.
I rilievi fotografici ed il campionamento distruttivo sono avvenuti tra l’autunno del 2010 e la primavera del 2011 (Regione del Veneto-Magistrato alle Ac-que di Venezia-Thetis-SELC, 2012).
Il campionamento tradizionale di tipo distruttivo, eseguito contemporaneamente a quello fotografico, ha previsto il grattaggio di 3 quadrati di 50 cm di lato, utilizzando martello, scalpello e prelevando il materiale con una sorbona; per i dettagli si rimanda a Miotti et al. (2014).
In fase di analisi delle immagini, dopo una prima valutazione, sono state se-lezionate, per ogni affioramento, almeno 10 immagini fotografiche di qualità ottimale (per trasparenza, profondità di campo, nitidezza, piano di ripresa a distanza costante) per il riconoscimento della componente macrozoobentonica. L’analisi degli scatti fotografici è avvenuta tramite un software di analisi delle immagini che, per le forme coloniali, ha permesso di calcolare la copertura dei taxa rispetto all’area di analisi, in accordo con le metodiche utilizzate per l’ana-lisi dei campioni mediante grattaggi (Miotti et al., 2014).
Per il riconoscimento tassonomico del macrozoobenthos in fase di post-proces-sing, si è fatto riferimento anche a schede riassuntive compilate direttamente in immersione dagli operatori e a collezioni di laboratorio.
L’analisi dei gradienti e della similarità tra le comuinità zoobentoniche degli af-fioramenti è stata eseguita mediante la Cluster Analysis (similarità di Bray-Curtis e dati trasformati con la radice quarta; Clarke & Warwick, 2001), la Multi-Dimensional Scaling (MDS) ed il calcolo degli indicatori: numero di taxa (S), copertura (Cop) e indice di diversità di Shannon (Shannon & Weaver, 1949).
Mediante il test ANOSIM si è verificata la possibilità di poter eseguire le analisi successive, utilizzando i valori medi derivati dall’analisi dei singoli scatti foto-grafici. I test, applicati attraverso l’analisi dell’indice di similarità di Bray–Cur-tis, hanno evidenziato una buona rappresentatività delle repliche fotografiche per ogni singolo affioramento (R=0,503; P=0,001).
Risultati e discussione
Nella tabella 2 sono riportati i valori di copertura media (%) per affioramento dei taxa determinati nelle fotografie, identificati a seguito delle analisi delle im-magini; essi hanno un range di variazione elevato, compreso tra il 10% dell’af-fioramento LA1-003 e il 65% della tegnùa di CAORLE, assestandosi media-mente su un valore del 28%.
I taxa che presentano i valori di copertura totale (somma delle coperture di tutti gli affioramenti) più elevati sono, per i Poriferi, Tedania anhelans e Cliona viridis e, per gli Ascidiacei coloniali, Aplidium conicum e Polycitor adriaticus.
Tab. 2. Valori medi di copertura per taxa, totale e per gruppo tassonomico.
| Abate | Pivetta | Lastre | LA1-003 | MA1-003 | Motta Sile | Cerniotta | Tartaruga | Caorle | Picco | Panetton | Piave | |
| Thetya spp. | 3.5 | 0 | 0 | 0 | 0 | 0.5 | 0 | 0 | 0 | 0.3 | 0.5 | 0 |
| Ircinia sp. | 0.6 | 3.4 | 0 | 0 | 6.5 | 0 | 0 | 1.4 | 4.9 | 0 | 0.1 | 0 |
| Aplysina aerophoba | 0.3 | 2.7 | 2.2 | 0.5 | 6.1 | 0.2 | 4.3 | 1.8 | 4.2 | 0 | 0 | 0 |
| Tedania anhelans | 0.3 | 0.4 | 6.5 | 0 | 19.9 | 2.8 | 2.5 | 19.4 | 0 | 0 | 1.5 | 0.8 |
| Cliona celata | 1.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cliona viridis | 1.7 | 7 | 2.5 | 1.4 | 0 | 12.3 | 3.1 | 0.9 | 17.4 | 0.9 | 4.7 | 11.3 |
| Porifero ind. | 0.8 | 0 | 0.9 | 1.5 | 0 | 1.2 | 0 | 0 | 1.2 | 2.5 | 3 | 1.8 |
| Halicondria sp. | 2.8 | 1.4 | 0 | 0 | 8.7 | 0 | 0 | 0 | 10.7 | 0 | 0 | 0 |
| Sarcotragus spinosulus | 0 | 7.6 | 0 | 0 | 1.3 | 0 | 0 | 0 | 1.6 | 0 | 0 | 0 |
| Aplidium tabarquensis | 0 | 0 | 0 | 0 | 0 | 0 | 0.3 | 0.3 | 0 | 0 | 0.3 | 0 |
| Aplidium conicum | 0 | 0.6 | 2.3 | 0 | 0 | 0 | 0 | 1.9 | 0 | 7.7 | 4.8 | 10.1 |
| Polycitor adriaticus | 0.1 | 8.6 | 0.4 | 0 | 0 | 4.1 | 7.4 | 2.1 | 0 | 3.8 | 3.2 | 3.9 |
| Clavelina sp. | 0.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chondrosia reniformis | 0 | 0 | 9.3 | 0.5 | 10.5 | 3 | 0 | 0 | 11.4 | 0 | 0 | 0 |
| Geodia cydonium | 0 | 0 | 0.5 | 0 | 0 | 1.4 | 0 | 0 | 0 | 0 | 0 | 0 |
| Dysidea spp. | 0 | 0 | 1.2 | 6.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 |
| Cystodytes dellechiajei | 0 | 0 | 0 | 0 | 0 | 0 | 0.6 | 0 | 0 | 0 | 0.5 | 0 |
| Schizobrachiella sanguinea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9.5 | 0 | 0 | 0 |
| Cladocora caespitosa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3.9 | 0 | 0 | 0 |
| Copertura totale | 12.3 | 31.7 | 25.8 | 10.3 | 53 | 25.5 | 18.2 | 27.8 | 64.8 | 15.2 | 18.7 | 27.9 |
| S | 10 | 8 | 9 | 5 | 6 | 8 | 6 | 7 | 9 | 5 | 10 | 5 |
| Porifera | 11.5 | 22.5 | 23.1 | 10.3 | 53 | 21.4 | 9.9 | 23.5 | 51.4 | 3.7 | 9.9 | 13.9 |
| Ascidiacea | 0.8 | 9.2 | 2.7 | 0 | 0 | 4.1 | 8.3 | 4.3 | 0 | 11.5 | 8.8 | 14 |
| Anthozoa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3.9 | 0 | 0 | 0 |
| Bryozoa | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9.5 | 0 | 0 | 0 |
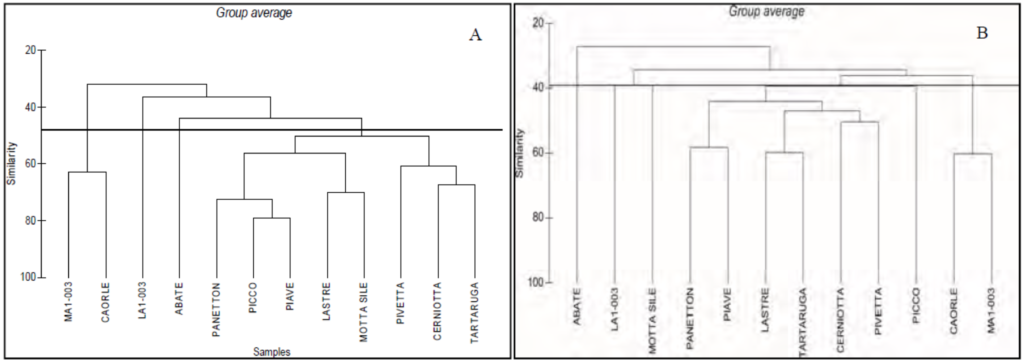
La Cluster Analysis, eseguita sui valori medi di copertura dei taxa identificati, evidenzia, alla similarità del 44%, quattro principali gruppi di affioramenti (Fig. 1A). Il gruppo con il maggior numero di affioramenti (MOTTA SILE, PICCO, PANETTON, PIAVE, PIVETTA, TARTARUGA, LASTRE e CERNIOTTA) include strutture rocciose situate in posizione intermedia tra la costa e il mare aperto (5,8-15 km), mentre l’affioramento ABATE, situato ad una distanza di oltre 28 km dalla costa, costituisce un raggruppamento a se stante.
Gli altri due raggruppamenti, quello degli affioramenti MA1-003 e CAORLE e quello costituito dal solo affioramento LA1-003, pur trovandosi in prossimità della costa (1-3 km), risultano però relativamente distanti geograficamente fra loro e si separano sulla base della maggiore biodiversità (MA1-003 e CAORLE) e dei bassi valori di biodiversità e copertura (LA1-003).
iotti et al., 2014).
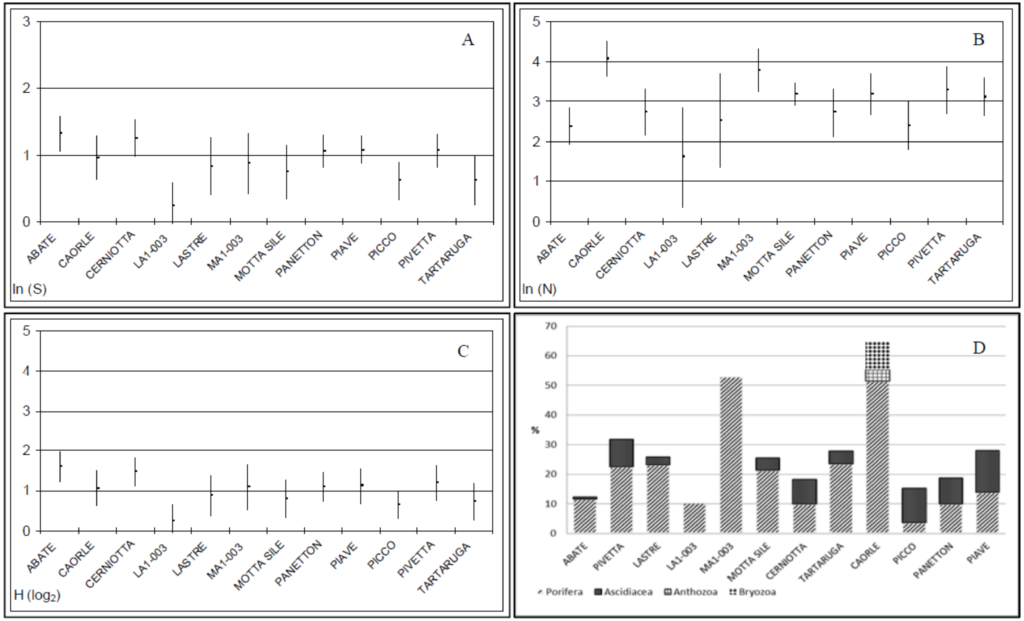
Il confronto tra i principali indicatori ecologici (S, Cop e H’), ricavati dai dati di copertura, evidenzia un grado più elevato di diversità della comunità zooben-tonica dell’affioramento ABATE, mentre le tegnùe CAORLE e MA1-003 spic-cano per copertura di forme coloniali (Fig. 2A, B); l’affioramento dove sono stati registrati i valori più bassi di tutti gli indicatori è l’affioramento LA1-003. Il confronto tra valori di copertura dei principali gruppi tassonomici (Porifera, Ascidiacea, Anthozoa e Bryozoa) evidenzia come negli affioramenti vicini alla costa (LA1-003, MA1-003 e CAORLE) non sia stata registrata la presenza di ascidiacei che abbondano, invece, in quelli intermedi (es. PIVETTA, PICCO, PANETTON, PIAVE) e sono presenti più al largo (ABATE) (Fig. 2D). Gli antozoi e i briozoi sono stati rilevati solo nell’affioramento CAORLE rispet-tivamente con le specie: Cladocora caespitosa e Schizobrachiella sanguinea. Per quanto riguarda S. sanguinea, una sua maggiore abbondanza in affioramenti di aree costiere era già stata osservata nell’ambito di altri lavori (Molin et al., 2010) e potrebbe essere legata al fatto che alcuni briozoi per la loro crescita necessitano di un certo grado di idrodinamismo e tassi di sedimentazione (Co-cito, 2009).
L’analisi di correlazione di Spearman condotta sui valori degli indicatori S, Cop e H’ e i parametri ambientali (coordinate, profondità e distanza dalla costa), ha evidenziato una correlazione altamente significativa tra il numero di taxa e il gradiente legato alla longitudine (p<0.001); le tegnùe localizzate nella parte orientale del golfo di Venezia presentano, infatti, un numero di taxa maggiore rispetto a quelle della costa occidentale. Dall’analisi di correlazione sono risultati significativi anche i confronti tra il numero di taxa negli affioramenti e la loro distanza dalla costa e profondità (p<0.01), mentre le coperture totali diminuiscono allontanandosi dalla costa e nelle tegnùe più profonde, evidenziando quindi una correlazione inversa (p< 0.05). Anche il confronto tra indice di Shannon e profondità/distanza dalla costa è risultato molto significativo (p<0.005).
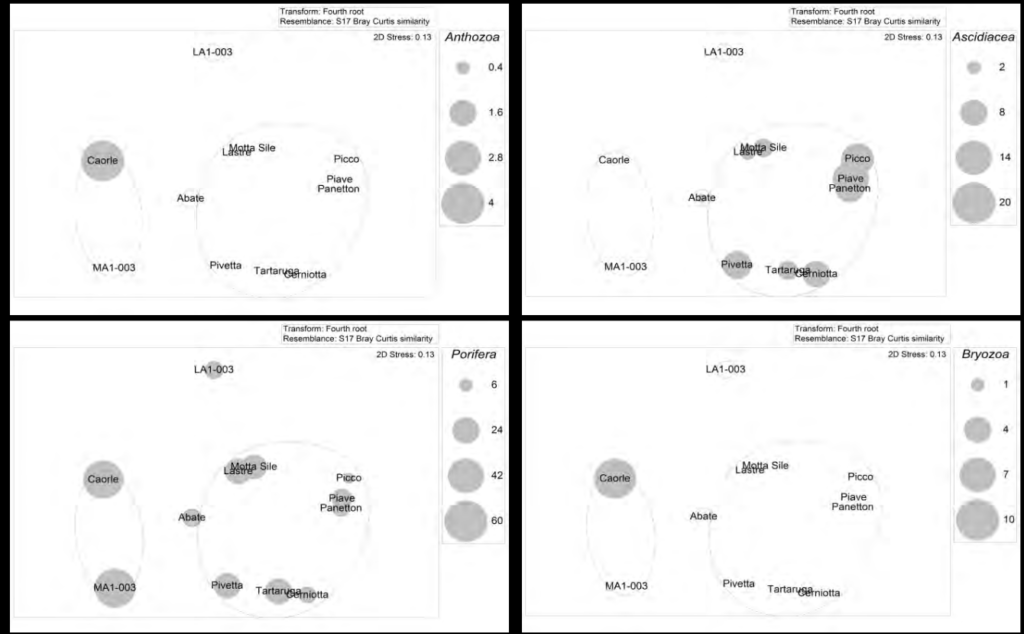
L’analisi MDS della distribuzione dei tre gruppi di taxa coloniali mostra come briozoi e antozoi (Schizobrachiella sanguinea e Cladocora caespitosa) hanno un’elevata copertura solo nell’affioramento CAORLE (Figure 3A e 3D) mentre i poriferi hanno una distribuzione più omogenea e sono presenti in tutti gli affior-manti, anche se con coperture maggiori nel gruppo CAORLE-MALAMOCCO (Fig. 3C).
Gli ascidiacei, nel loro complesso, sembrano in grado di caratterizzare meglio le comunità degli affioramenti posti a distanza intermedia dalla costa (Fig. 3B); in tutti gli affioramenti più vicini alla costa (1-3 km), questo gruppo è risultato assente (CAORLE, MA1-003, LA1-003), mentre abbonda in quelli intermedi tra la costa e il largo (5,8-15 km) per diminuire nuovamente al largo (oltre i 28 km) (ABATE).
Tab. 3. Matrice di correlazione di Spearman tra parametri ambientali e indici di comunità (in grassetto le relazioni statisticamente significative con P<0.05).
| R rs | S | Cop | H |
| Est | 0.601* | 147 | 0.490** |
| Nord | -168 | 294 | -280 |
| Prof. | 458 | -399 | 0.493** |
| Dist. cost | 462 | -399 | 0.483** |
*p<0.001; **p<0.005
Il confronto tra i risultati del presente lavoro, basato su una metodologia di indagine fotografica (non distruttiva), con quelli del campionamento mediante grattaggio (distruttivo) eseguito negli stessi siti (tre repliche di 50 x 50 cm e ana-lisi al microscopio della macrofauna) (Miotti et al., 2014) evidenzia coerenza nella ripartizione degli affioramenti e nell’andamento degli indicatori.
Ad un livello di similarità paragonabile (circa 40%), la cluster analysis delle due metodiche (Fig. 1) conferma sia lo schema di aggregazione degli affioramenti in 4 gruppi, sia l’aggregazione secondo il gradiente costa-largo, con la sola va-riazione dell’affioramento MOTTA SILE, che cambia di raggruppamento in ragione della sua posizione geografica “intermedia” tra la costa e il largo (Fig. 3). Per la metodica distruttiva (grattaggi), la comunità zoobentonica di questa tegnùa è più simile a quella dell’affioramento LA1-003 vicino alla costa presso la bocca di porto di San Nicolò del Lido, mentre per la metodica non distruttiva (fotografica) risulta simile al raggruppamento delle tegnùe poste ad una distan-za intermedia dalla costa.
Le analisi degli indicatori di comunità calcolati con la metodica fotografica han-no evidenziato andamenti comparabili a quelli riscontrati nei campioni raccolti con metodica distruttiva (Miotti et al., 2014), soprattutto per le coperture tota-li. Tra le due metodiche, invece, appaiono molto meno comparabili il numero di taxa e l’indice di Shannon, che risentono maggiormente dell’influenza delle specie criptiche non rilevabili dall’analisi fotografica (Tabella 4).
Per quanto concerne la biodiversità, l’analisi dei dati ricavati dalle fotografie è stata in grado di evidenziare comunque la presenza di gradienti, già osservati nell’ambito di precedenti studi condotti dagli autori (Molin et al., 2010; Curiel et al., 2012; 2014), che associano una composizione specifica più ricca e diffe-renziata agli affioramenti localizzati più ad est e più lontani dalla costa.
Nel complesso i dati di comunità confermano gli scarsi valori di copertura e numero di taxa dell’affioramento vicino alla bocca di Lido LA1-003 che risente della vicinanza della bocca lagunare e l’elevato valore di biodiversità della te-gnùa ABATE, la più lontana dalla costa.
| Tegùa | Metodo fotografico | Metodo distruttivo | ||
| N. taxa | Indice Shannon | N. taxa | Indice Shannon | |
| ABATE | 10 | 1,6 | 16 | 2 |
| CAORLE | 9 | 1,1 | 25 | 2,8 |
| CERNIOTTA | 6 | 1,5 | 11 | 1,6 |
| LA1-003 | 5 | 0,3 | 11 | 1,3 |
| LASTRE | 9 | 0,9 | 10 | 1 |
| MA1-003 | 6 | 1,1 | 16 | 1,8 |
| MOTTA SILE | 8 | 0,8 | 13 | 1,8 |
| PANETTON | 10 | 1,1 | 9 | 1,3 |
| PIAVE | 5 | 1,1 | 9 | 1 |
| PICCO | 5 | 0,6 | 9 | 1,5 |
| PIVETTA | 8 | 1,2 | 10 | 1 |
| TARTARUGA | 7 | 0,7 | 13 | 0,7 |
Conclusioni
I risultati delle indagini sulle comunità biologiche ottenuti con la metodica fo-tografica sono in linea con quanto emerso dalle indagini condotte con il pre-lievo degli organismi mediante il grattaggio e l’analisi succesiva al microscopio. La metodologia fotografica ha evidentemente dei limiti, intrinsechi al tipo di approccio analitico, tuttavia lo sviluppo della tecnologia digitale permette di procedere al campionamento di aree molto estese e coglie la necessità di trovare nuove metodiche di indagini non invasive, specie nelle aree protette o nei siti Natura 2000. Tale necessità ne ha favorito un forte utilizzo nel corso dell’ultimo decennio in tutto il Nord Adriatico (Molin et al., 2003; 2006c; 2008; Curiel et al., 2017;Ponti& Mastrototaro, 2006; Ponti et al., 2006; 2011;Regione delVeneto-Magistrato alle Acque di Venezia-Thetis-SELC, 2012; Regione del Veneto-CNR-ISMAR, 2014).
A fronte di un minor dettaglio tassonomico, i vantaggi di questo metodo sono evidenti e consistono in un impatto pressochè nullo sulla comunità bentonica, nella velocità e facilità di esecuzione, soprattutto nella fase di campionamento, che permette all’operatore subacqueo di estendere l’area di indagine o di ese-guire un campionamento rappresentativo anche a profondità significative con tempi di permanenza al fondo ridotti.
Dall’esame di due recenti lavori che hanno indagato la comunità zoobentoni-ca mediante scatti fotografici (Regione del Veneto-Magistrato alle Acque di Venezia-Thetis-SELC, 2012; Regione del Veneto-CNR- ISMAR, 2014) si rileva come con questa metodica sia possibile giungere al riconoscimento dello zoobenthos a livello di genere o di specie per circa 30-40 organismi.
Il livello di definizione tassonomica raggiungibile per la componente zoobento-nica dall’esame delle immagini fotografiche risulta ben superiore a quello per la componente macroalghe (Curiel et al., 2017), più critica sia per le dimensioni millimetriche di molte specie, sia per la definizione anche del solo genere, ren-dendo più opportuna un’analisi di questa comunità per gruppi funzionali (turf, incrostanti, erette) piuttosto che ad un livello tassonomico disomogeneo (es. specie, genere, ordine, classe, forme indet., ecc.).
A questi vantaggi, si aggiungono poi i minori costi operativi sia di campo che di analisi di laboratorio e la maggiore sicurezza degli operatori subacquei, soprat-tutto quando si interviene a profondità superiori a 20-30 m, sino alla soglia di una immersione tecnica (oltre 40 m).
Se si soppesano vantaggi e svantaggi, è chiaro che il metodo fotografico rap-presenta uno strumento oggi essenziale nei programmi di monitoraggio delle comunità bentoniche in aree protette o di alto valore ambientale; esso fornisce l’opportunità di pianificare attività di monitoraggio su ampia scala di questi particolari habitat, tra cui le tegnùe del Nord Adriatico, ed è in grado di rilevare le macroevidenze che possono fungere da early warning di eventuali forme di degrado di questi habitat.
Ringraziamenti
Si ringrazia il Ministero delle Infrastrutture e dei Trasporti – Magistrato alle Acque di Venezia (ora Provveditorato Interregionale alle OO. PP. del Veneto – Trentino Alto Adige – Friuli Venezia Giulia) tramite il Concessionario Consorzio Venezia Nuova per aver concesso l’utilizzo dei dati dello studio Op. 499 – Intervento di valorizzazione delle aree costiere prospicienti le bocche di porto della Laguna di Venezia. Creazione di zone di tutela biologica e marina in accordo con la Re-gione del Veneto. Monitoraggio delle caratteristiche ambientali e morfologiche delle tegnùe naturali collocate nei diversi ambiti di intervento in prossimità delle bocche di porto.
Bibliografia
Aa.Vv. (2010) – Le tegnùe dell’Alto Adriatico: valorizzazione della risorsa marina attraverso lo studio di aree di pregio ambientale. ARPAV: 52-61.
Adriatic sea forum (2016) – Adriatic Sea Tourism Report 2016 EDITION.
Bohnsack A.J. (1979) – Photographic quantitative sampling of hard-bottom benthic communities. Bull. Mar. Sci., 29(2): 242-252.
Casellato S., Stefanon S. (2008) – Coralligenous habitat in the northern Adriatic Sea: an overview. Mar. Ecol., 29: 321-341.
Clarke K.R., Warwick R.M. (2001) – Change in marine communities: an approach to statistical analysis and interpretation. 2nd Ed., PRIMER-E, Playmouth, UK: 172 pp.
Christi H. (1980) – Methods for ecological monitoring: biological interactions in a rocky subtidal community. Helgolandr Meeresunters, 33: 473-483.
CNA (2015) – Dinamiche e prospettive di mercato della filiera nautica del diporto. Cna Produzione Nazionale, pp. 120.
Cocito S. (2009) – Le biocostruzioni a briozoi. Biol. Mar. Mediterr., 16 (1): 19-30.
Curiel D., Falace A., Vinko B., Kaleb S., Solidoro C., Ballesteros E. (2012) – Species composition and spatial variability of macroalgal assemblages on biogenic reefs in the northern Adriatic. Sea. Bot. Mar., 55(6): 625-638.
Curiel D., Miotti C., Checchin E., Rismondo A, Cerasuolo C., Kaleb S., Falace A. (2014) – Biodiversità macroalgale e gradienti ecologici degli affioramenti rocciosi del litorale veneto. Boll. Mus. St. Nat. Venezia, 65: 5-21.
Curiel D., Molin E. (2010) – Comunità fitobentoniche di substrato solido. In: AA.VV., Le tegnùe dell’Alto Adriatico: valorizzazione della risorsa marina attraverso lo studio di aree di pregio ambientale. ARPAV: 62-79.
Curiel D., Miotti C., Checchin E., Rismondo A., Molin E. (2017) – Analisi delle macroalghe dei substrati rocciosi (tegnùe) del Nord Adriatico mediante rilievi fotografici e confronto con una metodica distruttiva. Lavori Soc. Ve. Sc. Nat., 42: 89-101.
Fedra K., Machan R. (1979) – A Self-Contained Underwater Time-Lapse Camera for in situ Long-Term Observations. Mar. Biol., 55: 239-246.
Gabriele M., Bellot A., Gallotti D., Brunetti R. (1999) – Sublittoral hard substrate communities of the northern Adriatic Sea. Cah. Biol. Mar., 40: 65-76.
Gage, J.D., Tyler, P.A. (1991) – Deep sea biology: a natural history of organisms at the deep sea floor. Cambridge University Press, Cambridge.
Gambi M.C., Dappiano M. (2003) – Mediterranean marine benthos: a manual of methods for its sampling and study. Biol. Mar. Medit., 11 (Suppl. 1), pp. 604 .
Lampitt, R.S., Billet, D.S.M. & Rice, A.L. (1986) – Biomass of the invertebrate mega-benthos from 500 to 4100 m in the Northeast Atlantic. Mar. Biol., 93: 69-81.
Miotti C., Checchin E., Curiel D., Rismondo A., Cerasuolo C., Molin E. (2014) –
Variazioni nelle comunità macrozoobentoniche di affioramenti rocciosi (tegnùe) del litorale veneto lungo un gradiente costa-mare. Boll. Mus. St. Nat. Venezia, 65: 47-65.
Mizzan L. (1992) – Malacocenosi e faune associate in due stazioni altoadriatiche a sub-strati solidi. Boll. Mus. civ. St. Nat. Venezia, 41: 7-54.
Mizzan L. (1995) – Le “Tegnùe”. Substrati solidi naturali del litorale veneziano: Poten-zialità e prospettive. ASAP, Venezia, 46 pp.
Molin E., Gabriele M., Brunetti R. (2003) – Further news on hard substrate communi-ties of the northern Adriatic Sea with data on growth and reproduction in Polycitor adriaticus (von Drasche, 1883). Boll. Mus. civ St. Nat. Venezia,54: 19-28.
Molin E., Maggiore F., Zanella M. (2006a) – Stime di biomassa di Tubularia crocea (Agassiz, 1862) mediante monitoraggio fotografico in un’area a barriere artificiali nel Nord Adriatico. Biol. Mar. Medit., 13 (1): 737-740.
Molin E., Gomiero M. (2006b) – Stime di biomassa di Mytilus galloprovincialis (LA-MARCK, 1819) e Styela plicata (LESUEUR, 1823) mediante monitoraggio fotografico in un’area a barriere artificiali nel Nord Adriatico. Biol. Mar. Medit., 13 (2):196-197.
Molin E., Gomiero M., Zanella M. (2006c) – Monitoraggio fotografico della comunità bentonica nel campo sperimentale. Il Campo Sperimentale in mare: prime esperienze nel veneto relative a elevazioni del fondale con materiale inerte. Quad. ARPAV ISBN 88-7504-104-0.
Molin E., Berton A. (2007) – A new approach to define the ecological status of North Adriatic hard substrate benthic communities of rocky outcrops. In: Pérez-Ruzafa A.,
Hoffmann E., Boncoeur J., Garcìa-Charton J.A., Salas F., Sorensen T.K., Vestergaard O. (eds.), European Symposium on Marine Protected Areas as a Tool for Fisheries Management and Ecosystem Conservation.Murcia(Spain), 25-28september: 242-244.
Molin E., Bocci M., Picone M., Penna G., Zanovello G. (2008) – Analisi fotografica del megabenthos in tre affioramenti rocciosi (tegnùe) del golfo di Venezia (Nord Adriatico). Biol. Mar. Medit., 15 (1): 276-277.
Molin E., Soccorso C., Bon D. (2009) – Stime di biomassa di Haliclona (Reniera) me-diterranea Griessinger, 1971 mediante monitoraggio fotografico in un’area a barriereartificiali nel Nord Adriatico e stato di colonizzazione del reef artificiale. Boll. Mus. civ. St. nat. Venezia,59: 19-26.
Molin E., Pessa G., Rismondo A. (2010) – Comunità macrozoobentonica di substrato solido. In: Aa.Vv. (2010. Le tegnùe dell’Alto Adriatico: valorizzazione della risorsa marina attraverso lo studio di aree di pregio ambientale. ARPAV: 52-61.
Ponti M., Mastrototaro F. (2006) – Distribuzione dei popolamenti ad ascidie sui fondali rocciosi (Tegnùe) al largo di Chioggia (Venezia). Biol. Mar. Medit., 13 (1): 621-624 Ponti M., Tumedei M., Colosio F., Abbiati M. (2006) – Distribuzione dei popolamenti epibentonici sui fondali rocciosi (Tegnùe) al largo di Chioggia (Venezia). Biol. Mar. Medit., 13(1): 625-628.
Ponti M., F. Fava, Abbiati M. (2011) – Spatio-temporal variability of epibenthic as-semblages on subtidal biogenic reefs in the northern Adriatic Sea. Mar. Biol., 158: 1447-1459.
Pronzato, R. (1996) – Underwater photographic techniques for field-research in shal-low marine environments. In European Artificial Reef Research, ed. Jensen, A.C., Proceedings of the 1st EARRN Conference, Ancona, Italy, March, 337-346.
Regione del Veneto, CNR-ISMAR (2014) – Attività di monitoraggio riguardanti la distribuzione ed estensione degli habitat bentonici e pelagici. Substrati biogenici (Tegnùe). 70 pp.
Regione del Veneto – Magistrato Alle Acque (ora Provveditorato Interregionale alle OO. PP. del Veneto – Trentino Alto Adige – Friuli Venezia Giulia) – Thetis – SELC (2012) – Intervento di valorizzazione delle aree costiere prospicienti le boc-che di porto della laguna di Venezia. Creazione di zone di Tutela Biologica e Marina in accordo con la Regione del Veneto. OP 499. 1. Rapporto finale delle attività di ricerca in mare, 169 pp.
Shannon C.E., Weaver W. (1949) – The mathematical theory of communication. Urbana, Univ. Illinois Press.
Shears N.T., Russel C., Babcock C. (2003) – Continuing trophic cascade effects after 25 years of no take marine reserve protection. Mar. Ecol. Prog. Ser., 246: 1-16.
Weinberg, S. (1978) – The minimal area problem in invertebrate communities of Medi-terranean rocky substrata. Mar. Biol., 49: 33-40.