Riassunto
Vengono elencate le specie di Lepidotteri reperite nel corso delle ricerche condotte negli anni 2016-2017 nella Riserva Integrale di Bosco Nordio. È di particolare interesse tassonomico un esemplare del genere Oegoconia sp., che probabilmente rappresenta una specie inedita; sono nuove per la fauna dell’Italia continentale: Neurothaumasia tenuipennella Gaedike 2011, Oegoconia ceres Sutter 2007, Cosmiopterix crassicervicella (Chrétien, 1896), Coccidiphila danilevskyi Sinev, 1997, Psamathocrita argentella Pierce & Metcalfe, 1942; Cydia rymarczyki Varenne & Nel 2013, Cydia (Kenneliola) sammuti Diakonoff 1986, Merulempista brucella (Studiger, 1879). Sono nuove per la regione Veneto: Dryadaula heindeli Gaedike & Scholz 1998, Paradoxus osyridellus Millière 1869, Megacraspedus balneariellus Chrétien 1907, Pammene giganteana (Peyerimhoff, 1863). Il ritrovamento di esemplari di sesso femminile di Cydia (Kenneliola) sammuti permette di descrivere la femmina fino ad ora sconosciuta. Una nuova forma di Argyresthia abdominalis Zeller, 1839 è illustrata.
Abstract
Contribution to the knowledge of the Lepidoptera of the Integral Regional Natural Reserve “Bosco Nordio”
The species of Lepidoptera found during the fieldwork in the years 2016-2017 in the Integral Reserve of Bosco Nordio are reported. A specimen of the genus Oegoconia is of particular taxonomic interest and probably belongs to an undescribed species; new to the fauna of continental Italy are the following species: Neurothaumasia tenuipennella Gaedike 2011, Oegoconia ceres Sutter 2007, Cosmiopterix crassicervicella (Crétien, 1896), Coccidiphila danilevskyi Sinev, 1997, Psamathocrita argentella Pierce & Metcalfe, 1942; Cydia rymarczyki Varenne & Nel, 2013, Cydia (Kenneliola) sammuti Diakonoff 1986, Merulempista brucella (Studiger, 1879). They are new to the Veneto region:Dryadaula heindeli Gaedike & Scholz 1998, Paradoxus osyridellus Millière 1869, Megacraspedus balneariellus Chrétien 1907, Pammene giganteana (Peyerimhoff, 1863). The finding of female specimens of Cydia (Kenneliola) sammuti allows to describe the female until unknown. A new form of Argyresthia abdominalis Zeller, 1839 is illustrated.
Introduzione
L’entomofauna del Bosco Nordio è poco nota. Le conoscenze attuali sono principalmente quelle su Ortotteri e i Coleotteri (Canestrelli, 1980; Contarini, 1997) e sui Lepidotteri Geometridi (Flamigni et al., 2005). Scopo del presente lavoro è documentare la comunità dei Lepidotteri, un gruppo faunistico la cui presenza è legata alla vegetazione essendo il regime alimentare dei bruchi essenzialmente fitofago. Lo studio delle famiglie di tutto l’ordine Lepidoptera permette di avere informazioni sulle cenosi delle diverse tipologie di habitat. I Lepidotteri sono inoltre facilmente campionabili e sono efficaci bioindicatori della conservazione dell’ambiente come evidenziano numerosi lavori pubblicati per la regione Veneto (Huemer & Triberti, 2004; Huemer & Morandini, 2009; Triberti et al., 2017, 2018; Timossi, 2009, 2013, 2014).
Area di studio
Il Bosco Nordio, tra i siti Natura 2000 della Regione Veneto, ha il codice IT3250032 e fa parte del raggruppamento 5B, Biotopi litoranei e sistemi dunali (Buffa G., Lasen C., 2010). Le superfici dichiarate SIC e ZPS, 157 Ha, sono coincidenti. Il Bosco Nordio è situato sul sistema dunale più antico del litorale altoadriatico, tra il fiume Adige e il fiume Po. L’antica foresta era nota già in epoca preromana con il nome di Cerreto o Fosson (Simonella, 2006). Situato in località Sant’Anna nel comune di Chioggia, il Bosco Nordio è una delle poche testimonianze dell’ampia fascia boscosa che un tempo caratterizzava il litorale Veneto. Ne sono altri esempi il Bosco della Mesola, a sud, il Bosco di Valgrande di Bibione a nord. L’origine del sistema dunale di Bosco Nordio è dovuta all’azione del Po dell’Adige e del mare che negli ultimi 4000 anni hanno trasportato e distribuito enormi volumi di sedimenti (Ghirelli, 2001).
Da oltre quarant’anni l’area è una Riserva Naturale Regionale Integrale gestita da Veneto Agricoltura (atto di istituzione del 1971). La storia della foresta è legata alla famiglia Nordio, da cui il nome del bosco, che dal 1565 ai primi anni del ’900 sfruttò la tenuta a scopo agricolo disboscando la lecceta originaria. A fine ’800 venne introdotto il pino domestico (Pinus pinea) probabilmente per ricavare legna e pinoli data la bassa redditività agricola (Ghirelli, 2001). Allo stato attuale l‘ente gestore persegue l’obiettivo del recupero della vegetazione originaria introducendo essenze arboree e arbustive autoctone, agevolato dal deperimento spontaneo della pineta che, anche per motivi fitopatologici, negli ultimi decenni è in regresso. Nel corso degli ultimi anni sono state ripristinate alcune depressioni umide creando specchi d’acqua alimentati dalla falda superficiale dell’Adige. Negli anni ’60 venne costruita una strada che congiunge la statale Romea all’Adige e divide in due settori l’area della Riserva. Il presente lavoro riguarda lo studio del settore meridionale che, oltre ad essere la superficie maggiore, è oggetto delle azioni di ripristino da parte dell’ente gestore. In questo settore inoltre dal 1964 al 2000 era presente il Daino, specie introdotta a scopo venatorio, che è stata definitivamente rimossa in tempi recenti per i danni che causava allo sviluppo naturale della vegetazione (Ghirelli, 2001).
Caratteristiche ambientali, flora e vegetazione (fig. 1)
L’assetto geomorfologico del Bosco Nordio è quello di una serie di cordoni dunali paralleli alla linea di costa intervallati da depressioni umide. Nella parte più alta delle dune la vegetazione è costituita da cenosi a leccio (Quecus ilex), cod. Habitat 9340, mentre nelle aree interdunali umide è presente il querceto caducifolio a farnia (Quercus robur). La pineta monospecifica (cod. Habitat 2270*) è ridotta a pochi alberi e per lo più deperienti. Il sottobosco della lecceta, che è dominante, è composto da specie termofile a distribuzione mediterranea quali il pungitopo (Ruscus aculeatus), l’asparago dei boschi (Asparagus acutifolius), la robbia selvatica (Rubia peregrina), Clematis flammula, il lilatro (Phyllirea angustifolia), l’orniello (Fraxinus ornus). Dal punto di vista fitosociologico le cenosi a leccio del litorale altoadriatico sono extrazonali e differiscono da quelle mediterranee per la presenza di elementi eurasiatici quali ligustro (Ligustrum vulgare), biancospino (Crataegus monogyna) e sanguinella (Cornus sanguinea).
Nelle radure e ai margini dei sentieri sono presenti lembi di duna grigia (cod. Habitat 2130*) costituiti da depositi sabbiosi e sabbioso-ghiaiosi stabilizzati e non raggiunti dall’acqua del mare (Buffa G., Lasen C, 2010; (Ghirelli, 2001).
Nell’adriatico settentrionale (nei litorali di Friuli Venezia Giulia, Veneto e Emilia Romagna) la vegetazione delle dune grigie viene riferita all’associazione Tortulo- Scabiosetum Pignatti, 1953 che è considerata endemica (Buffa G., Lasen C, 2010; http://vnr.unipg.it/habitat/index.jsp) delle coste nord adriatiche comprese tra la foce del Tagliamento a est e il litorale di Ravenna, poco oltre la foce del fiume Po, a ovest.
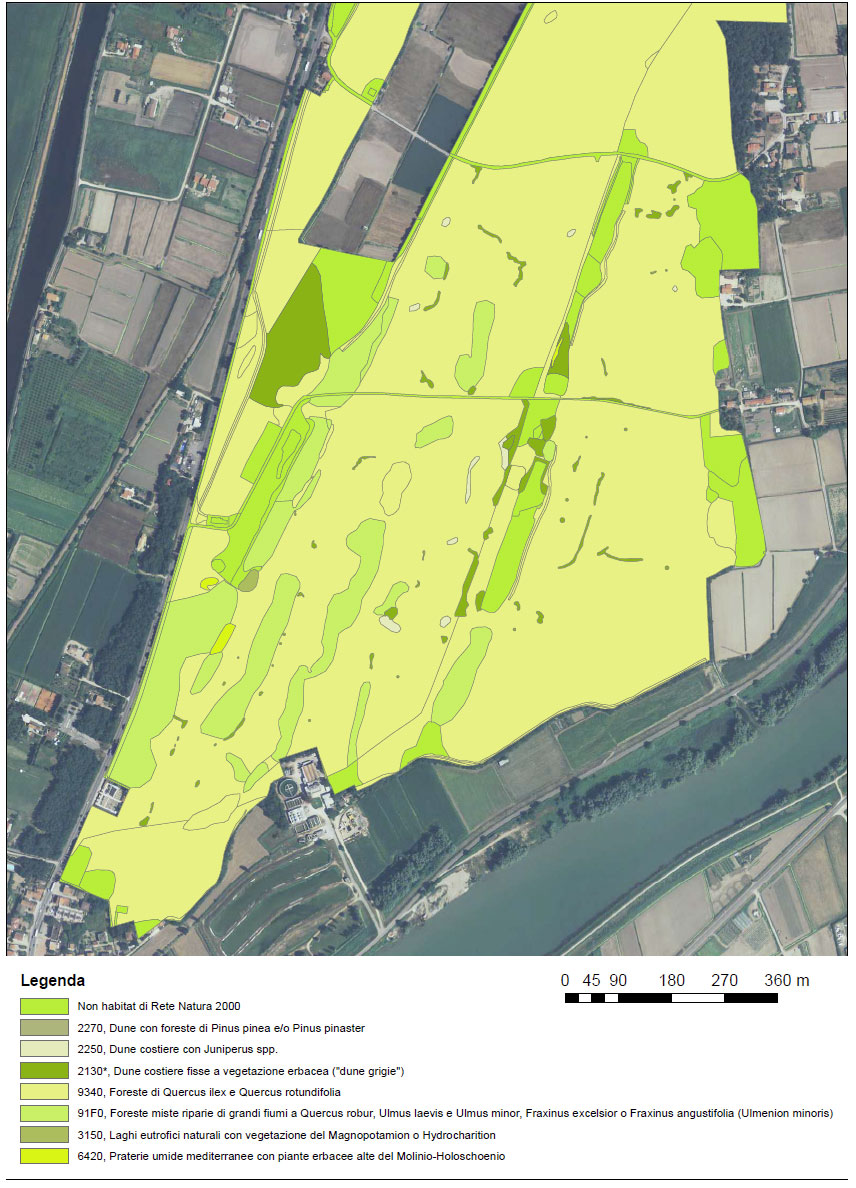
Questo habitat è strettamente correlato al bosco extrazonale di leccio; al Bosco Nordio, è isolato dall’Adige dai cordoni dunali mobili litoranei ad Ammophila arenaria (cod. Habitat 2120). Negli spazi aperti xerici si trova anche il ginepro (Juniperus spp.) e che qui va considerato specie guida dell’habitat 2250* che nella regione Veneto è estremamente localizzato. Nelle depressioni umide, ricostruite con interventi appropriati e preservate dall’interramento, l’Habitat presente può essere interpretato come 6420 – Praterie umide mediterranee con piante erbacee alte del Molinio–Holoschoenion. Sono presenti a contorno dei laghetti l’immancabile cannuccia di palude (Phragmites australis), il giunco (Juncus litoralis) e in acqua la ninfea (Nymphaea alba).
Entomofauna del Bosco Nordio
Il Bosco Nordio è stato oggetto raramente di ricerche specifiche sull’entomofauna.
I pochi contributi pubblicati sono quelli di Canestrelli (1980) sugli Ortotteri, con 16 specie, e di Contarini (1997) sui Coleotteri, con 91 specie.
Qualche altra segnalazione faunistica è presente in pubblicazioni generali sulla laguna di Venezia (Ratti, 1986, 1991; Zangheri, 1958). Più di recente Flamigni et al. (2005) pubblicano dati inediti di Geometridi, di cui vengono segnalate per l’area 30 specie. Per quanto riguarda il Bosco Nordio, il materiale studiato è stato raccolto dal prof. Sergio Zangheri e risale agli anni 1964-1972: i reperti sono conservati nelle collezioni del Dipartimento agronomia ambientale e produzioni vegetali (ora Dafnae) dell’Università di Padova. Alla lista di specie di Flamigni et al. (2005) segnalate per il Bosco Nordio vanno aggiunte le seguenti specie reperite nel corso delle ricerche: Eupithecia ericeata (Rambur, 1833), Cyclophora annularia (Fabricius, 1775), Scopula rubiginata (Hufnagel,1767), Idaea rusticata ([Denis & Schiffermüller], 1775), Idaea politaria (Hübner, [1799]), Rhodometra sacraria (Linnaeus, 1767).
Materiali e metodi
Le ricerche sono state condotte negli anni 2016-2017 da Aprile a Settembre: le date di raccolta figurano nella tabella 1. La metodologia di ricerca dei Lepidotteri è consolidata (Leraut, 2014; Parenti, 2000). Nella ricerca delle specie ad attività diurna si utilizza il retino entomologico per catturare a vista i lepidotteri che si alzano in volo, o si usa il retino da sfalcio per le specie che sostano sulle piante erbacee e gli arbusti. Per le specie ad attività notturna sono state utilizzate diverse fonti di luce di attrazione nel corso delle sessioni di monitoraggio.
Combinando lampade al sodio (Na, 160 W), tubi fluorescenti a gas inerti (luce attinica 20 W, di Wood 15 W) contemporaneamente, posti davanti a un telo bianco sorretto da appositi supporti, o all’interno di una tenda di tulle, è stato possibile raccogliere la maggior parte dei reperti (Parenti, 2000). I macrolepidotteri sono raccolti con flaconi in polipropilene con all’interno acetato di etile; i microlepidotteri studiati sono stati raccolti vivi in provette apposite in vetro e successivamente preparati in laboratorio secondo metodologie standardizzate (Parenti, 2000). È stata utilizzata anche una trappola automatica di raccolta.
Posizionata prima del tramonto, con accensione a sensore di luce crepuscolare, e con anestetico a base di ammoniaca, e stata lasciata in azione tutta la notte. Al mattino veniva ispezionato e raccolto il contenuto. Un laboratorio temporaneo di ricerca è stato allestito nella foresteria dell’ente di gestione sita nella riserva.
Le osservazioni in natura di farfalle diurne in volo sono state condotte catturandole con il retino e dopo l’identificazione gli esemplari sono stati rilasciati.
In collezione i reperti sono individuati con cartellino di colore verde chiaro con dicitura: I; Veneto, Ve, Chioggia, Bosco Nordio – habitat, Staz. n°. Tutto il materiale è raccolto dall’autore.
La preparazione degli apparati genitali si basa sulle le indicazioni di Robinson (1976) e per i Gelechiidae di Huemer & Karsholt (1999). Per alcuni esemplari si è preferita la conservazione degli apparati in glicerina con microcapsula spillata sotto l’esemplare al posto dell’usuale montaggio su vetrino.
La determinazione degli esemplari è dell’autore tranne che i Coleophoridae che sono stati determinati dallo specialista Giorgio Baldizzone.
Documentazione fotografica
Per le fotografie dei preparati microscopici è stato utilizzato un microscopio Nikon Eclipse E100 munito di microcamera Sony Color CCD 5.1 Mp TP 5100 con software X-Entry. Per la documentazioni fotografica è stata usata una Camera Nikon D200 + Nikon PB-5Bellows + Nikon 105mm f/2.8 Micro – NIKKO, illuminazione con 3 spot light e software Combine ZP per lo stacking.
Dna barcoding
La sequenza di codici a barre del DNA si basa su un segmento di 648 coppie di basi del gene COI mitocondriale (citocromo c ossidasi 1). Campioni di zampe metatoraciche disidratate sono stati inviati e sequenziati dal Canadian Centre for DNA Barcoding (CCDB, Biodiversity Institute of Ontario, University of Guelph e dalla compagnia AIM (Advanced Identification Method). Le sequenze di Megacraspedus balneariellus, Cosmopteryx crassicervicella e Argyresthia abdominalis sono state depositate in BoldSystem, così come l’immagine dell’esemplare, seguendo le indicazioni raccomandate (Ratnasingham & Hebert, 2007).

Stazioni di monitoraggio (fig. 2)
Le stazioni temporanee di rilevamento sono state poste nelle radure e in prossimità dello stagno grande.
Stazione n° 1; N 45°.118298, E 12°.260587 – radura prati aridi.
Stazione n° 2; N 45° 121527, E 12° 262094 – radura prati mesofili.
Stazione n° 3: N 45° 120891, E 12° 256137 – stagno grande prati umidi.
Stazione n° 4: N 45° 119209, E 12°25776 – depressione umida in bosco.
Nel corso della ricerca diurna e al tramonto, è stato percorso il transetto che unisce la stazione 1 alla stazione 2 percorrendo la radura arida fino allo stagno e fino al prato mesofilo. Inoltre è stata percorsa la strada forestale di giorno e sono state ispezionate le radure adiacenti alla stessa.
Lista delle specie
La lista di specie è ordinata secondo il catalogo dei lepidotteri europei di Karsholt & Razowski (1996). Il codice numerico è quello del catalogo: per le specie descritte successivamente alla pubblicazione del catalogo segue una lettera al numero del genere. Se il genere è nuovo per la fauna europea viene attribuito il numero dell’ultimo genere elencato con l’aggiunta della lettera.
L’inquadramento delle famiglie segue l’ordinamento di Nieukerken et al. (2011). Seguono le colonne: genere e specie, autore, sesso, stadio di sviluppo della specie reperita (bruco o adulto), n° degli esemplari reperiti, numero della stazione di monitoraggio, data di cattura o di osservazione. Segue la descrizione del regime alimentare del bruco tratta dalla letteratura conosciuta e da osservazioni sul campo.
Nella colonna successive vi sono alcune informazioni sulla autoecologia delle specie studiate attribuendo le specie a delle categorie ecologiche. Le categorie sono le seguenti: per la preferenza per il grado di umidità specie di ambiente igrofilo, mesofilo, xerofilo; sulla base delle piante ospiti se è specie di ambienti forestali (specificando se conifere o latifoglie), arbusteti, prati, ecotoni, sistemi dunali, paludi (di acqua dolce o salmastre), ubiquista; l’acronimo S evidenzia le specie stenoecie per l’habitat presente al Bosco Nordio: la relazione tra la specie e l’habitat è desunta verificando che la pianta, o le piante, alimentare del bruco sia nella lista delle piante presenti nella combinazione fisionomica di riferimento per gli habitat presenti nell’area di monitoraggio. Se la pianta alimentare non è nota l’attribuzione è sulla base dalle informazioni di bibliografia e da osservazioni personali. Viene riportato se la specie è importante per conservazione dell’entomofauna saproxilica (Campanaro et al., 2011); segue la colonna che specifica se è stata catturata alla luce (lux) o di giorno e in cui sono riportate anche annotazioni di altra natura.
Specie di interesse tassonomico
Oegoconia sp. (Symmocidae) fig. 3.
Reperti: stazione 1; 2.VI.2016, ♀, prep. mic. 1597 (tav I, fig. a, b, c).
Note: Sutter (2007) descrive nuove specie del genere Oegoconia di ambiente mediterraneo e pubblica una chiave dicotomica per il riconoscimento delle specie Europee del genere. In accordo con la trattazione tassonomica di Gozmany (2008, pag. 435) l’esemplare reperito ha l’ostium, il signum e l’VIII tergite che non sono riconducibili ad alcuna specie descritta: In particolare l’ostium è largo e aperto, il signum ovale e piegato al centro con una spina apicale ad angolo ottuso.
Apertura alare 11 mm, II° e III° segmento dei palpi labiali neri con apice bianco. Nella trattazione del genere Oegoconia Sutter (2007) figura una serie di reperti dei Balcani che non sono attribuiti a nessuna specie nota; l’esemplare studiato sembra una specie diversa. Sarà indispensabile reperire anche i maschi per una diagnosi accurata.
Biologia: sconosciuta: le specie del genere Oegoconia si nutrono per lo più di detriti vegetali e foglie morte nella lettiera.
Distribuzione: nota solo del Bosco Nordio.
Habitat: foreste di Quercus ilex.
Specie nuove per la fauna dell’Italia continentale
Neurothaumasia tenuipennella Gaedike, 2011 (Tineidae) fig. 4.
Reperti: stazione 1, 27.VII.2016, 2 ♀ prep. mic. 1596 (tav I, fig. d).
Note: nuova per la fauna dell’Italia continentale! Gli esemplari raccolti corrispondono a quelli figurati da Gaedike (2011, fig. 1-2, 9; 2015, figs. 68).
Biologia: sconosciuta: si presume che il bruco viva nelle gallerie di Cerambycidae in cui si nutre di detrito e residui di cuticola larvale (Gaedike, 2015).
Distribuzione: Italia: Sardegna, Veneto. Europa: Romania, Grecia (Creta e Lesbo), Croazia (Krk), Spagna, Francia (Gaedike, 2015).
Habitat: foreste di Quercus ilex.
Oegoconia ceres Sutter, 2007 (Symmocidae) fig. 5.
Reperti: stazione 1, 2.VI.2016, ♀ prep. mic. 1584 (tav I, fig. e, f, g).
Note: specie descritta recentemente della Sardegna (Sutter, 2007): locus typicus Monte Arcosu (Ca), Sa Canna. È nuova per l’Italia continentale! Gozmany (2008) non la riporta nella lista di specie del genere Oegoconia perché la sua monografia è uscita postuma.
Biologia: le specie del genere Oegoconia si nutrono per lo più di detriti vegetali e foglie morte della lettiera.
Distribuzione: Italia: Sardegna, Veneto.
Habitat: foreste xeriche di Quercus.
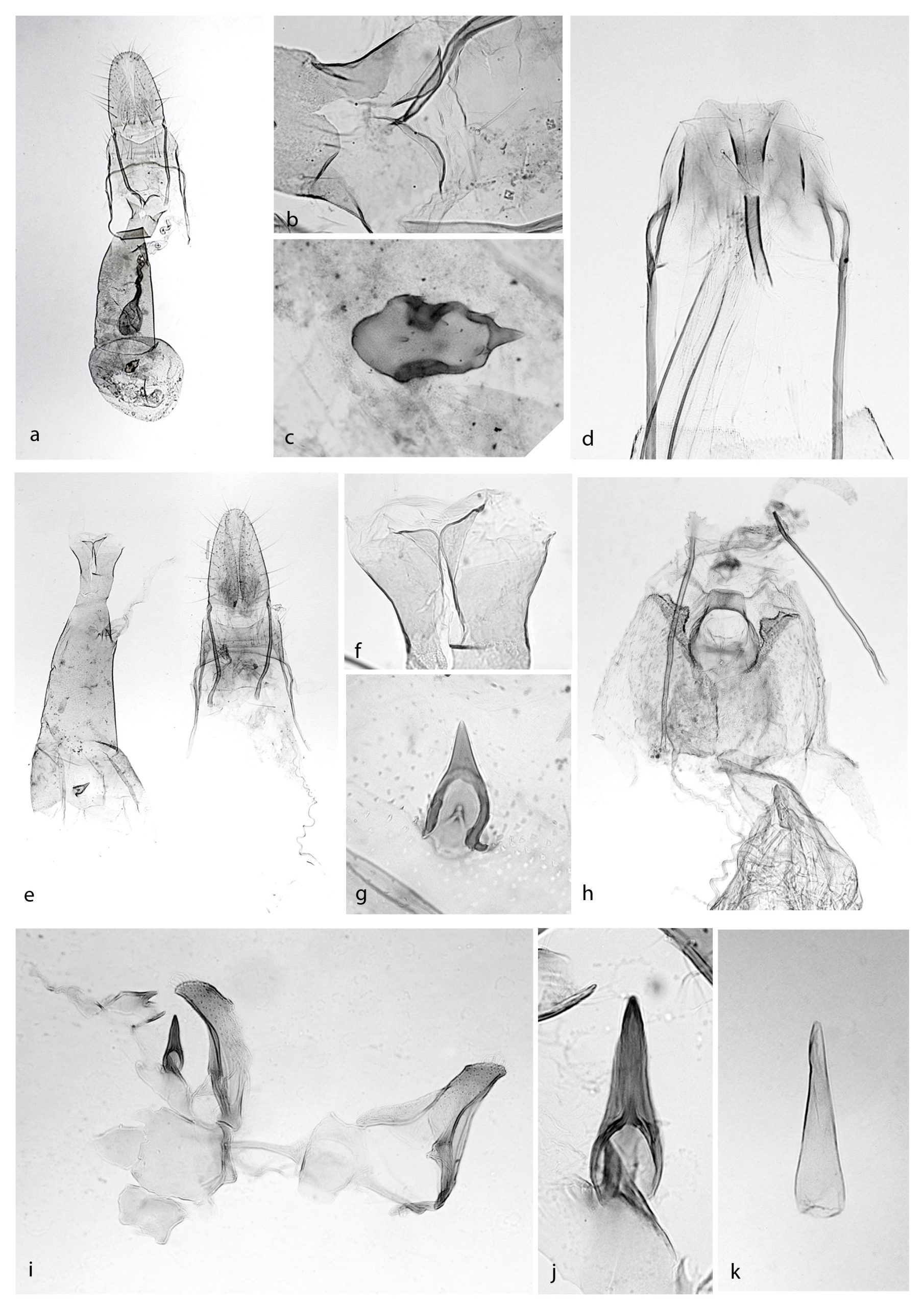
Oegoconia sp.:
a. genitali femminili,
b. hostium,
c. signum;
Neurothaumasia tenuipennella:
d. ostium;
Oegoconia ceres:
e. genitali femminili,
f. ostium,
g. signum;
Coccidiphila danilevskyi:
h. ostium;
Psamathocrita argentella:
i. valve genitali maschili,
h. uncus,
k. edeagus.

Fig 3 
Fig 4 
Fig 5 
Fig 6
Fig. 4. Neurothaumasia tenuipennella ♀ (foto D. Valloto).
Fig. 5. Oegoconia ceres ♀ (foto D. Valloto).
Fig. 6. Cosmopteryx crassicervicella ♀ (foto D. Valloto).
Cosmopteryx crassicervicella (Cretien, 1896), (Cosmopterigidae) fig. 6.
Reperti: stazione 1, 27.VII.2016, 4♀, prep mic. 1588, 1583 [BOLD id. AIM20012018_2] (fig. 7); idem, 4.IX.2016, ♀, prep. mic. 1577.
Note: nuova per la fauna italiana! Per la difficoltà di studio degli apparati copulatori femminili, in cui i caratteri diagnostici sono meno evidenti che nei maschi, e per la difficoltà nel distinguerla dalle congeneri, ho inviato le zampe posteriori di un esemplare per lo studio del DNA barcode. L’analisi genetica ha confermato la specie e l’analisi dei genitali femminili è stata comparata con quelli figurati da Kostner & Sinev (2003; fig. 76).
Biologia: pianta alimentare del bruco Cyperus spp. (Cyperaceae), (Kostner & Sinev, 2003).
Distribuzione: Italia: Veneto. Europa: Spagna, Portogallo, Grecia: dall’Asia minore al Nord Africa (Kostner & Sinev, 2003).
Habitat: prati umidi.
Coccidiphila danilevskyi Sinev, 1997 (Cosmopterigidae) fig. 8.
Reperti: stazione 1, 27.VII.2016, ♀ prep. mic. 1612 (tav. I, fig. h).
Note: nuova per la fauna italiana! Kosten & Sinev (2003) segnalano per l’Italia per il genere Coccidiphila le seguenti specie: C. gerasimovi Danilevsky, 1950 e C. ledereriella (Zeller, 1850). L’esemplare reperito è incompleto, ma comunque lo studio dei genitali femminili ne consente la determinazione. In particolare ostium e sterigma corrispondono al quelli figurati da Kosten & Sinev (2003; fig. 97).
Biologia: sconosciuta; le larve delle altre specie del genere Coccidiphila si nutrono di uova di Homoptera Coccoidea, di residui animali in nidi di lepidotteri e di ragni, di foglie morte, di galle e semi di leguminose.
Distribuzione: Italia: Veneto. Europa: Francia, Spagna, Marocco e Tunisia (Kosten & Sinev, 2003).
Habitat: forestale?.
immagine

Psamathocrita argentella Pierce & Metcalfe, 1942 (Gelechiidae) fig. 9.
Reperti: stazione 1, 2.VI.2016, ♂, prep. mic. 1517 (tav I, fig. i, j, k).
Note: specie nuova per l’Italia. Del genere Psamathocrita sono presenti in Europa tre specie: P. argentella Pierce & Metcalfe, 1942, P. dalmatinella Huemer & Tokar, 2000 e P. osseella (Stainton, 1861). Da quando Huemer et. al. (2004) segnalarono la presenza di esemplari di questo genere nel litorale altoadriatico, la questione dell’identità specifica è rimasta dubbia. Effettivamente, la specie è molto rara e sono pochi gli esemplari da me raccolti in anni di ricerche negli ambienti salmastri della Laguna di Venezia. Sono altresì pochi gli esemplari raccolti e studiati anche da altri specialisti. La difficoltà di studio degli apparati genitali maschili e la scarsità di esemplari mi hanno indotto a confrontarmi con altri studiosi per una diagnosi sicura.
Biologia: pianta alimentare del bruco Agropyrum pungens.
Distribuzione: Italia: conosciuta del litorale alto adriatico: Bosco Nordio, Valle Millecampi (dati inediti), Valle Vecchia in provincia di Venezia. Europa: Regno Unito, Francia;
Habitat: ambienti litoranei: barene(?).
Immagine

Fig 8 
Fig 9
Fig. 9. Psamathocrita argentella ♂ (foto D. Valloto).
Cydia rymarczyki Varenne & Nel, 2013 (Tortricidae) fig. 10.
Reperti: stazione 1, 4.IX.2016, ♂ prep. mic. 1678 (tav. II, fig. l).
Note: nuova per la fauna italiana! Habitus e genitali del solo esemplare reperito corrispondono a quelli pubblicati da Varenne & Nel (2013, fig. a-b, 6, 9). La specie era nota solo della Francia.
Biologia: sconosciuta; ha fenologia tardiva e vola in un’unica generazione a fine estate. I bruchi delle specie congeneri si nutrono di frutti di Quercus, Corylus e Juglans.
Distribuzione: Italia: Veneto. Europa: Francia.
Habitat: foreste di Quercus ilex.
Cydia (Kenneliola) sammuti Diakonoff, 1986 (Tortricidae) fig. 11, 12, 13.
Reperti: stazione 1, 4.IX.2016, 2 ♂ prep. mic. 1669 (tav. II, fig. m), 1673; idem, ♀ prep. mic. 1674 (tav II, fig. n, o, p).
Note: il genere Cydia conta 78 specie in Europa (Razowski, 2003). Diakonoff (1986) descrive C. sammuti sulla base di un unico esemplare maschio di Malta e lo assegna al sottogenere Kenneliola. La femmina non era ancora nota.
Descrizione della femmina: la nomenclatura del disegno alare e delle parti dei genitali segue quella di Razowski (2003). Apertura alare 16 mm; capo: colore ocra con squame orientate in avanti alla fronte e a formare una cresta al vertice; antenne: filiformi con cilia corte e ricoperte di squame brune; palpi labiali: secondo segmento ocra, terzo segmento di colore bruno e lungo 1/2 del terzo; tegulae e torace colore ocra; ali anteriori: colore di fondo ocra-bruno nella metà basale, più scure che nel maschio, strisce costali rossastre, linee marginali dello speculum indistinte, linee interne allo speculum nere, frange bicolori; ali posteriori: grigio scuro con frange bicolori. Apparato genitale femminile: apofisi posteriori e anteriori delle stessa lunghezza, sterigma con postostial plate di forma tronco-conica, ostium bursae ampio e sclerotizzato, collicolum sinuoso lungo quasi fino al margine anteriore del settimo sternite, corpus bursae piriforme, presenti due piccoli signum conici, dritti con apice arrotondato. Razowski (2003) nella lista sistematica dei tortricidi europei colloca C. sammuti successivamente a C. exquisitana (Rebel, 1889) che è la specie più affine; i caratteri differenziali diagnostici dei genitali femminili sono: apofisi anteriori lunghe quanto quelle posteriori, collicolum sinuoso e lungo, mentre in C. exquisitana le apofisi sono di diversa lunghezza e il collicolum è molto corto: i signum sono grandi in C. sammuti mentre in C. exquisitana sono minuti.
Biologia sconosciuta: nella descrizione del locus typicus Diakonoff cita la presenza di Pinus halepensis e Lonicera implexa: la flora del Bosco Nordio annovera tra le piante mediterranee in comune con quelle citate Lonicera etrusca che ritengo possa essere la pianta alimentare del bruco. L’esemplare di Malta descritto da Diakonoff è stato raccolto in Giugno mentre al Bosco Nordio la specie vola in Settembre.
Distribuzione: Italia: Veneto. Europa: Malta.
Habitat: boschi, boscaglie e siepi mediterranee.
Merulempista brucella (Staudiger, 1879), (Pyralidae) fig. 14, 15.
Reperti: stazione 1; 11.V.2016, ♀, prep mic. 1638 (tav II, fig. q).
Note: nuova per l’talia! Storia del taxon. Staudiger nella descrizione originale di Pempelia brucella la differenzia dalla congenere cingilella (Zeller, 1846) per l’apertura alare maggiore e per le ali anteriori sfumate di marrone-rosso. Il locus typicus citato per gli esemplari inviatigli da Mann è “Brussa” (= Bursa), Turchia.
Ragonot (1893) degrada P. brucella a varietà di P. cingilella; Roesler accetta la diagnosi di Ragonot e istituisce il genere Merulempista (Roesler, 1967) per cingilella (Zeller, 1846) e descrive Merulempista cingilella hartigi sulla base di un maschio di Policoro in Basilicata raccolto da Hartig F. (= M. c. harteri Roes. 1967,
immagine

Fig 10 
Fig 11 
Fig 12 
Fig 13 
Fig 14 
Fig 15
Fig. 11. Cydia (Kenneliola) sammuti ♂ (foto D. Valloto).
Fig. 12. Cydia (Kenneliola) sammuti ♂ (foto D. Valloto).
Fig. 13. Cydia (Kenneliola) sammuti ♀ (foto D. Valloto).
Fig. 14. Merulempista brucella ♀ (foto D. Valloto).
Fig. 15. Merulempista brucella ♂ (foto D. Valloto).
Asselbergs 1997, lapsus calami). Asselbergs (1997, fig 5, 8; 9, 12) descrive, M. wolschrjini dell’Olanda, e rivaluta M. brucella come specie sulla base dei caratteri degli apparati copulatori maschili e femminili. Leraut (2002, fig. 21) invece accetta la diagnosi di Ragonot (1893) e considera M. brucella sinonimo di M. cingilella; successivamente Leraut, (2014) mette in sinonimia M. wolschrjini con M. jucundella (Joannis, 1911). Per finire Nuss et al. (2019) confermano le tesi di Leraut (2002, 2014). Nel database pubblico Boldsystem (Ratnasingham & Hebert, 2007) un esemplare della Grecia di M. brucella è distinto anche dal punto di vista molecolare da M. cingilella.
Diagnosi: l’esemplare raccolto al Bosco Nordio è una femmina: lo studio degli apparati genitali concorda con la diagnosi di Asselbergs (1997, fig. 12) in cui il carattere discriminante più evidente è la forma convessa dell’antrum. Allo studio dei genitali dell’esemplare raccolto è stato affiancato lo studio comparativo dei genitali di esemplari provenienti dal litorale veneto raccolti dal collega Lucio Morin: Caorle, Brussa, loc. Valle Vecchia, 5.V.2000, ♂ prep. Mic. 1675 (tav. II, fig. r, s, t); idem, ♀; coll. privata Lucio Morin, Ronchi dei Legionari (Go).
La diagnosi comparativa di Asselbergs (1997, fig. 16) è confermata dallo studio dei genitali di un esemplare ♀ di M. cingilella di altra località: Gorizia, Grado, loc. Bonifica della Vittoria, 24.V.2001; legit e det. Lucio Morin; prep. mic. 1976 (fig. 16); coll. privata Lucio Morin, Ronchi dei Legionari (Go). M. brucella non è presente nella checklist on- line della fauna d’Italia (Stoch, 2003) e in P.E.S.I. (Pesi, 2019).
Biologia: sconosciuta: Leraut (2014) riporta per le specie del genere Merulempista come piante alimentari Tamarix spp. e Myricaria germanica e Salicornia (?).
Distribuzione: Italia: Veneto. Europa: Grecia, Bulgaria, Turchia.
Habitat: paludi di acqua dolce.
Specie nuove per il Veneto
Dryadaula heindeli Gaedike & Scholz, 1998, fig. 17.
Reperti: bosco, 11.V.2016, ♂ prep. mic. 1609 (tav. II, fig. u).
Note: nuova per il Veneto! Reperita di giorno in volo nel sottobosco.
Biologia: larve su funghi a mensola (Bjerkandera adusta) e del legno morto (Auricularia mesenterica).
Distribuzione: Italia: Veneto, Friuli Venezia Giulia. Europa: Germania, Svizzera, Francia, Spagna (Gaedike, 2015).
Habitat: foreste di latifoglie e conifere.
Paradoxus osyridellus Millière, 1869 (Yponomeutidae) fig. 18. Reperti: stazione 1, 26.V.2017, ♂.
Note: nuova per il Veneto!
Biologia: larva su Osyris alba, pianta tra le più significative specie mediterranee presenti al Bosco Nordio.
Distribuzione: Italia: Veneto, Venezia Giulia (Deutcsh, 2008). Europa: Spagna, Portogallo, Francia, Slovenia, Croazia, Montenegro, Grecia.
Habitat: prati aridi e dune grigie, macchia mediterranea.
Megacraspedus balneariellus Chretien, 1907 (Gelechidae) fig. 19, 20. Reperti: stazione 1, 11.V.2016, 3 ♂, prep mic. 1512, 1516 (tav. II, fig. v), [Bold id.n°LEASS623-17]; idem, 26.V.2017, 4 ♂.
Note: nella recente revisione del genere Megacraspedus, Huemer & Karsholt (2018) includono M. balneariellus nel gruppo di specie di M. fallax. Fino alla scoperta della popolazione di Bosco Nordio non era nota per il Veneto. Lo studio del DNA barcode di uno degli esemplari reperiti ha confermato la determinazione che appariva difficoltosa con lo studio dei soli caratteri morfologici degli apparati genitali maschili. Le femmine sono brachittere e non sono ancora state trovate.
Biologia: sconosciuta. Solo di poche specie del genere Megacraspedus è nota la pianta alimentare del bruco: si presume si nutra su Poaceae come altre congeneri.
Distribuzione: Italia: Veneto, Toscana: 3♀, prov. Grosseto, Marina di Albarese, Maremma, 19.VI.1981, leg. M. & E. Arenberger (Huemer & Karsholt, 2018).
Europa: Croazia, Francia, Spagna.
Habitat: biotopi litoranei: duna grigia (?).
Pammene giganteana (Peyerimhoff, 1863) Reperti: stazione 1, 13.IV.2016, 2 ♂, prep. Mic. 1578. Note: prima segnalazione per il Veneto!
Biologia: larva in galle di Hymenoptera (Andricus, Biorrhiza, Cynips) su Quercus robur, Castanea sativa. Sverna come pupa (Razowski, 2003).
Distribuzione: Italia: Veneto, Trentino – Alto Adige, Marche , Lazio, Molise, Puglia e Sicilia (Trematerra, 2003). Europa: Gran Bretagna, Francia, Germania, Svizzera, Austria, Slovenia, Bosnia, Serbia, Croazia, Romania, Bulgaria, Rep. Ceca.
Habitat: foreste mesofile.

Fig 16 
Fig 17 
Fig 18 
Fig 19 
Fig 20

Fig. 17. Dryadaula heindeli ♂ (foto D. Valloto).
Fig. 18. Paradoxus osyridellus ♂ (foto D. Valloto).
Fig. 19. Megacraspedus balneariellus ♂ (foto M. Uliana).
Fig. 20. M. balneariellus ♂, particolare del capo (foto M. Uliana)
Nuova forma cromatica di una specie
Argyresthia abdominalis Zeller, 1839
(Yponomeutidae) fig. 21, 22. Reperti: stazione 1, 26.V.2017, 3 ♂, prep. mic. 1571, 1573, 1574 [BOLD id. AIMSEQ20012018_3] (fig. 23).
Note: illustro uno degli esemplari reperiti per la particolarità del disegno delle ali anteriori: al Bosco Nordio tutti gli esemplari raccolti hanno un disegno delle ali incompleto rispetto alla forma nominale: in particolare le fasce ocra sono molto ridotte; la fascia mediana bianca ampia che inizia dalla base dell’ala raggiunge la stria ocra che da 2/3 della costa curva verso il tornus: manca del tutto la stria ocra curva che dalla metà del dorsum prosegue fino a 2/3 del centro dell’ala, inoltre le strie apicali sono evanescenti e la macchia nera apicale è assente.
Questa forma cromatica non mi risulta descritta. Nel nord Europa è nota una forma con ridotte fasce bianche (Wocke, 1884; f. decimella). A. abdominalis si può confondere con A. friulii (Huemer, 2009) che è descritta e conosciuta esclusivamente del Monte Canin in Friuli.
Biologia: la larva si nutre di aghi di Juniperus.
Distribuzione: Italia: nota del nord e del sud della penisola, in Veneto è nota del litorale di Caorle, Valle vecchia (Huemer & Morandini, 2008).
Habitat: è quello della pianta alimentare, nota per un notevole adattamento a diversi ambienti aridi dalle alte quote agli ambienti mediterranei.

Fig 21 
Fig 22
Fig. 22. A. abdominalis ♂, particolare ala destra (foto D. Valloto).

Analisi faunistica
Nella tabella 1 sono elencate 196 specie appartenenti a 36 famiglie. Se si sommano le specie di Geometridi pubblicate da Flamigni et al. (2005) e non reperite nel corso delle ricerche la fauna di Lepidotteri conta 221 specie. La Famiglia con il maggiore numero di specie, sul totale delle specie note, è quella dei Geometridae con 36 specie, Tortricidae con 23 specie, segue la famiglia Pyralidae con 19 specie, seguita dalle Famiglie Erebidae e Noctuidae con 18 specie ciascuna, Crambidae con 15 specie e Gelechiidae con 14. A livello di Superfamiglie quella con il maggior numero di specie è la Geometroidea a cui segue la Noctuoidea con 41 specie, seguita dalla Superfamiglia Gelechioidea con 35 specie. Le specie reperite nel corso della ricerca sono presenti per il 78% nella stazione 1, per il 23% nella stazione 3, il 22% nella stazione 2 e il 5% nella stazione 4. Per quanto riguarda il grado di umidità il 44% delle specie è presente in ambienti mesofili, il 31% in ambienti xerici, il 10,7 % in ambienti igrofili.
Gli ambienti in cui si ha la più alta biodiversità sono quelli forestali con il 41% delle specie studiate, di cui il 2,5% legato a conifere; il 32% delle specie frequenta spazi aperti come i prati e il 4% è legato a situazioni ecotonali; 4% è inoltre anche il contingente di specie presente in sistemi dunali. Una percentuale del 6% delle specie presenti è caratteristica di ambienti paludicoli: di acqua dolce il 4%, di ambienti salmastri il 2%. Sono ubiquiste eurecie il 10% delle specie reperite. Sono 1,5% le specie degli arbusteti e sono sinantropiche 1% delle specie reperite. Se prendiamo in considerazione gli habitat caratteristici del Bosco Nordio e le specie stenoecie per questi habitat, l’8% delle specie reperite è presente nell’habitat 91F0, il 6% nell’ habitat 9340, il 5% nell’habitat 3150, il 3,5% nell’habitat 2130. All’habitat 6420 sono legate il 2,5% delle specie, il 2% all’habitat 2270 e il 1,5% al 2250. Sono specie saproxiliche il 2% delle specie. Il 32% delle specie reperite è stenoecia e ha esigenze alimentari e abiotiche ristrette.
Discussione
La ricerca su tutto l’ordine Lepidoptera ha permesso di raccogliere una notevole quantità di dati inediti per l’area di studio. Se dal punto di vista biogeografico la presenza di 8 specie nuove per la fauna italiana aumenta le conoscenze sulla biodiversità del nostro paese, a livello regionale si ha un’idea più precisa della unicità di alcuni ambienti relitti come il Bosco Nordio. Sono stati riscontrati elementi caratteristici per ogni ambiente. In particolare per gli ambienti della lecceta extrazonale del litorale sabbioso alto adriatico sono stenoecie: Stigmella suberivora, Ectoedemia haraldi, Acrocercops brongniardella, Paradoxus osyridellus, Oecogonia ceres, Acrobasis glaucella e le specie del genere Cydia e Cyclophora puppillaria; Stigmella suberivora, Ectoedemia haraldi e Acrobasis glaucella sono monofaghe su Quercus ilex, queste specie hanno distribuzione mediterranea.
Nel querceto delle depressioni interdunali umide si evidenziano specie saproxiliche legate anche al grado di umidità maggiore come i tineidi Dryadaula heindeli, Nemapogon variatella, Neurothaumasia ankerella. La farnia è la pianta alimentare dei bruchi di Heliozela sericiella, Aleimma loeflingiana.
Ancylis laetana e Colobochyla salicalis sono anch’esse caratteristiche perché legate a Salix e Populus, piante presenti nella combinazione fisionomica di riferimento per l’habitat 91F0. Tra tutte le specie stenoecie per l’habitat delle dune grigie che è prioritario di conservazione, il Tortulo-Scabiosetum Pignatti, 1953, la specie più significativa è il crambide Platytes alpinella, la pianta alimentare del cui bruco è Tortula sp. I bruchi di Coleophora mareki e Thiodia trochilana si alimentano su diverse specie di Teucrium che sono associate ai muschi nella combinazione fisionomica di riferimento per l’Habitat 2130. Alla presenza di Juniperus, specie guida per l’habitat 2250 delle dune marittime mediterranee,
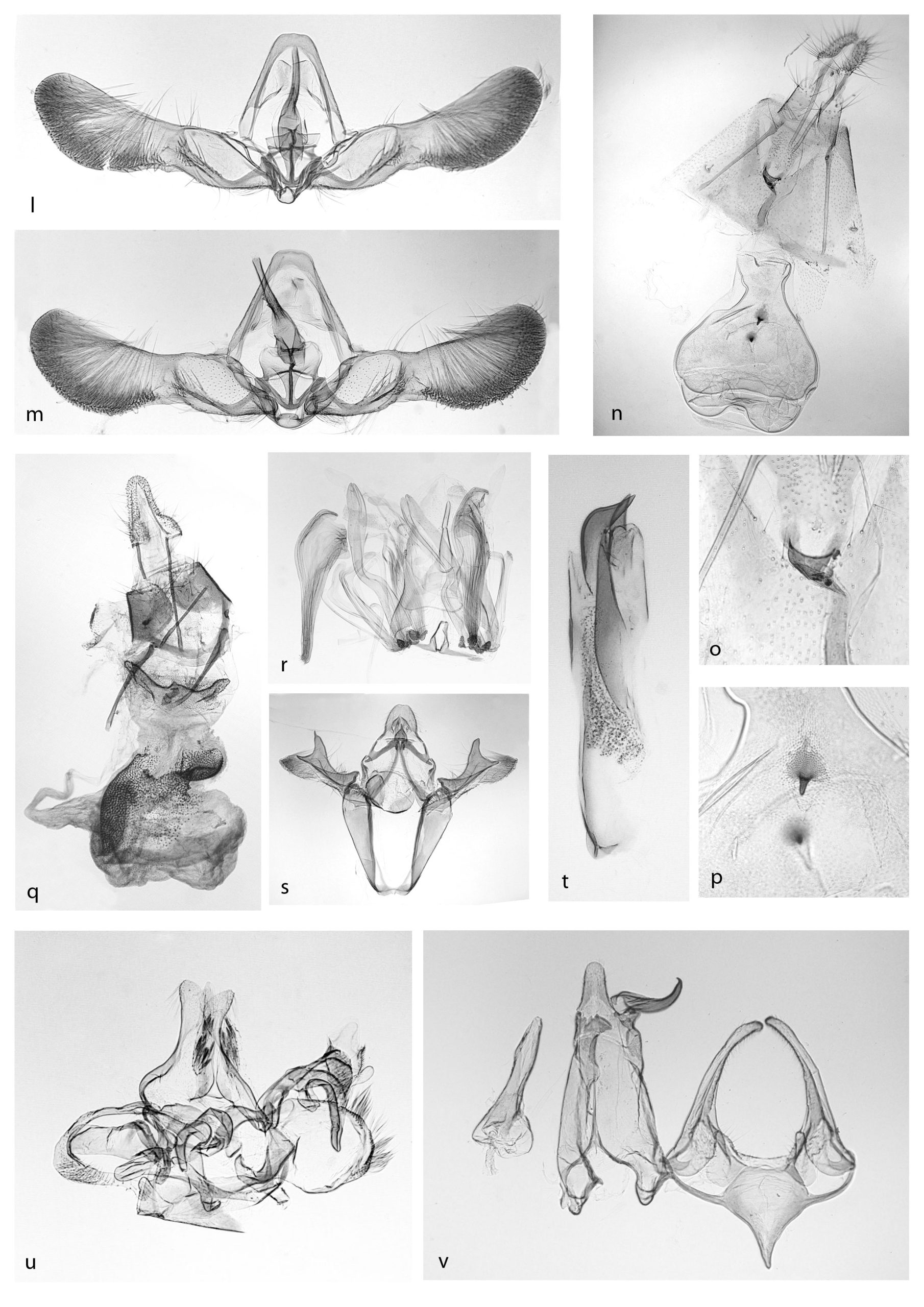
Cydia rymarczyki:
l genitali maschili;
Cydia (Kenneliola) sammuti:
m. genitali maschili,
n. genitali femminili,
o. ostium,
p. signum;
Merulempista brucella:
q. genitali femminili,
r. VIII° sternite maschile,
s. valve,
t. edeagus;
Dryadaula heindeli:
u. genitali maschili;
Megacraspedus balneariellus:
v. genitali maschili.
sono legate Argyresthia abdominalis e Eupithecia ericeata. Sono specie alloctone introdotte in Italia le seguenti specie: Coleophora texanella e Hyphantria cunea del Nord America, Tuta absoluta del Sud America, Duponchelia fovealis del Sud Africa.
Conclusioni
Ai fini della conservazione delle specie di Lepidotteri l’ente che gestisce la riserva integrale ha svolto nel passato e sta tuttora attuando strategie condivisibili e efficaci a questo scopo. In passato la rimozione del daino ha contribuito notevolmente alla rinaturalizzazione soprattutto della vegetazione delle dune e alla ripresa della lecceta. Così come l’estirpazione delle piante alloctone invasive.
Con il ripristino e l’ampliamento delle aree umide è conservata la biodiversità di quegli elementi igrofili caratteristici che con l’interramento delle pozze sarebbero scomparsi. La gestione delle foreste appare efficace, visto l’elevato numero di specie legate agli ecosistemi forestali sia della lecceta che del querceto igrofilo. Il rinvenimento di un’entità tassonomica incerta, o probabilmente nuova, suggerisce di continuare ad indagare questi ecosistemi per reperire altro materiale da studiare. A questo punto diventa fondamentale avviare ricerche anche nelle altre quercete extrazonali quali il Bosco della Mesola, a sud e il Bosco di Valgrande di Bibione a nord. La regione costiera del Veneto è altamente antropizzata. Sfruttata intensivamente a fini agricoli nell’entroterra, per il turismo nella fascia litoranea, è densamente urbanizzata e in pochi decenni ha perso molto della biodiversità che presentava prima dello sviluppo industriale del dopoguerra. Malgrado ciò le aree naturali relitte ancora presenti conservano specie vegetali e animali endemiche. Purtroppo questi biotopi, visto il contesto degradato in cui sono inseriti, sono destinati a perdere gradualmente la loro biodiversità se non verranno creati corridoi faunistici di connessione tra aree contigue che ne scongiurino l’isolamento. Per questo motivo la fauna del Bosco Nordio e i suoi habitat peculiari sono a rischio elevato di estinzione.
Ringraziamenti
Questo lavoro non sarebbe stato possibile senza la collaborazione di specialisti delle diverse famiglie di Lepidotteri. Ringrazio Peter Huemer (Gelechiidae) del Tiroler Landesmuseen-Betriebsgesellschaft m.b.H. (Innsbruck), Giorgio Baldizzone, Asti (Coleophoridae), Piero Giuseppe Varalda, Alessandria (Elachistidae), Paolo Triberti, Verona (Gracillaridae), Jacques Nel, La Ciotat (Tortricidae), Zdenko Tokàr, Michalovce (Gelechiidae). Per l’iconografia del presente lavoro ringrazio Davide Vallotto, Spinea e Marco Uliana del laboratorio di entomologia del Museo di Storia Naturale di Venezia Giancarlo Ligabue. Un ringraziamento a Federico Vianello e a Jacopo Richard di Veneto Agricoltura per l’incarico affidatomi. Mi sono avvalso della collaborazione di Giacomo Masato del Museo di Storia Naturale di Venezia Giancarlo Ligabue per la parte documentale.
Un ringraziamento ai soci della SVSN che mi hanno accompagnato sul campo: Enrico Bizio, Simone Bramuzzo e Davide Vallotto. Grazie a Jerome Morinière della compagnia AIM (Monaco). Si ringrazia il Canadian Centre for DNA Barcoding (Guelph, Canada), il cui lavoro di sequenziamento è stato permesso da finanziamenti del governo del Canada attraverso l’Ontario Genomics Institute. Ringrazio inoltre per il contributo alla stesura del manoscritto Alessandro Minelli, Giovanni Caniglia e il revisore anonimo.
Bibliografia
Asselbergs J., (1997) – Merulempista wolschrijni sp. nov. from the Netherlds and Merulempista brucella (Studiger) stat. nov. from Türkei (Lepidoptera, Pyralidae, Phycitinae). Ent. Ber. Amst. 57 (1): 8-13.
Buffa C., Lasen G. ( 2010) – Atlante dei siti Natura 2000 del Veneto. Regione del Veneto – Direzione Pianificazione Territoriale e Parchi. Venezia. pp 394.
Campanaro A., Bardiani M., Spada L., Carnevale L., Montalto F., Antonini G., Mason F. & Audisio P. (2011). Linee guida per il monitoraggio e la conservazione dell’entomofauna saproxilica. Verona: quaderni conservazione Habitat 6, Cierre Grafica.
Canestrelli P. (1980) – Gli ortotteri della riserva integrale di Bosco Nordio (Venezia). Ortotterofauna veneta (2° contributo). Lavori Soc. Ven. Scien. Nat. Vol. 5, pp. 3-8. Venezia.
Contarini E. (1997) – Osservazioni sulla coleotterofauna di un relitto di bosco termofilo della costa veneta: la R.N.I. di Bosco Nordio: Boll. Mus. Civ. St. Nat. di Venezia, vol. 47, pp. 75-94. Venezia.
Diakonoff A., (1986) – Two new Grapholini from Malta e Madeira (Lepidoptera, Totricidae). Nota lep. 9 (3-4): 170-174.
Deutcsh H. (2008) – Beiträge zur Lepidopterenfauna von Friaul Julisch Venetien (Norditalien). Teil 2: Provinz Gorizia und Trieste (Karst, Nordlichens Adriaticsches Küstengebiet). Contribution to knowledge of the Lepidoptera of Friuli Venezia Giulia (North Italy). Vol. 2: Province of Gorizia and Trieste (Karst, northern coast of Adriatic sea). Gortania, Atti Museo Friul. Di Storia Nat. 30:149-220.
Flamigni C., Zangheri S.,Rallo G., Uliana M., Gianti M. (2005) – Contributo alla conoscenza dei geometridi del Veneto: litorale, Colli Euganei e pianura circostante (Lepidoptera Geometridae). Boll. Mus. Civ. St. Nat. di Venezia, 56:113-135.
Gaedike R. (2015) – Tineidae I (Dryadaulinae, Hapsiferinae, Euplocaminae, Scardiinae, Nemapogoninae and Meessiinae). – In: Nuss, M., Karsholt, O. & P. Huemer [edit.]: Microlepidoptera of Europe 7: 1-308; Leiden & Boston (Brill).
Ghirelli L. (2001) – Indagini floristico-vegetazionali della Riserva Naturale di Bosco Nordio, pp. 90-98. In Cassol M. (2001). Guida alle riserve naturali in gestione a Veneto Agricoltura.
Gozmány L. (2008) – Microlepidoptera Palaearctica 13. Symmocidae, pp. 1-558. Goecke & Evers, Keltern.
Huemer P. (2010) – Argyresthia friulii sp. n., a new Species from the Julian Pre-alps, Northern Italy (Lepidoptera, Yponomeutidae). Gortania. Botanica, Zoologia, 31 (2009): 137-142.
Huemer P., Karsholt O. (1999) – Gelechiidae I (Gelechiinae: Teleiodini, Gelechiini). In Huemer, P., Karsholt, O. & Lyneborg, L. (Hrsg.): Microlepidoptera of Europe 3: 1-356. Apollo Books ed. Denmark.
Huemer P., Karsholt O. (2018) – Revision of the genus Megacraspedus Zeller, 1839, a challenging taxonomic tightrope of species delimitation (Lepidoptera, Gelechiidae). ZooKeys 800: 1-278.
Huemer P., Morandini C., Morin L. (2004) – New records of Lepidoptera for the italian fauna (Lepidoptera). Gortania, Atti Mus. Friu. di St. Nat., 26: 261-274, Udine.
Huemer P., Triberti P. (2004) – I Lepidotteri. In: Latella L. (ed.). Il Monte Pastello. Memorie del Muse Civico di Storia Naturale di Verona. 2 serie. Monografie naturalistiche 1: 177-208.
Huemer P, Morandini C. (2009) – Biodiversity of Lepidoptera within the area of Valle Vecchia (Caorle, Venezia) whit special regard to nature conservation aspect. Contributo alla conoscenza dei Lepidotteri dell’area di Valle Vecchia (Caorle, Venezia) con particolare riguardo agli aspetti conservazionistici. Gortania – Atti Museo Friul. Di Storia Nat., 30 (2008) pp. 221-254.
Karsholt O., Razowski J. (1996) – The Lepidoptera of Europe. A Distributional Checklist.Ed. Apollo Books, Stenstrup (DK).
Kostner J.C., Sinev S. Yu. (2003) – Momphidae, Batrachetidae, Stathmopodidae, Agonoxenidae, Cosmopterigidae, Chrysopeleiidae. In P. Huemer, O. Karsholt and Lyneborg (eds): Microlepidoptera of Europe 5: pp. 387.
Leraut P. (2002) – Contribution à l’étude de Phycitinae (Lepidoptera, Pyralidae). Nouv. Revue Ent. (N.S.), 19 (2):141-177.
Leraut P. (2014) – Moths of Europe vol IV. Pyralids 2. N. a. p. eds., France.
Nieukerken EJ van, Kaila L, Kitching IJ, Kristensen NP, Lees DC, Minet J, Mitter C, Mutanen M, Regier JC, Simonsen TJ, Wahlberg N, Yen S-H, Zahiri R, Adamski D, Baixeras J, Bartsch D, Bengtsson BÅ, Brown JW, Bucheli SR, Davis DR, Prins J De, PrinsW De, Epstein ME, Gentili-Poole P, Gielis C, Hättenschwiler P, Hausmann A, Holloway JD, Kallies A, Karsholt O, Kawahara A, Koster SJC, Kozlov M, Lafontaine JD, Lamas G, Landry J-F, Lee S, Nuss M, Park KT, Penz C, Rota J, Schmidt BC, Schintlmeister A, Sohn JC, Solis MA, Tarmann GM, Warren AD, Weller S, Yakovlev RV, Zolotuhin VV, Zwick A. (2011) – Order Lepidoptera Linnaeus, 1758. In: Zhang Z-Q (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 3148: 212-221.
Parenti U. (2000) – A Guide to the Microlepidoptera of Europe. Museo Regionale di Scienze Naturali di Torino, Guide I: 426 pp.
Ragonot E., (1893) – Monographie des Pycitinae et Galleriinae. In Romanoff, N. M., Memoires sur les lepidopteres. Mem. Lep. Rom. 7:1-658.
Ratnasingham S, Hebert PDN (2007) – BOLD: The Barcode of Life Data System (http://www. barcodinglife.org). Molecular Ecology Notes 7: 355-364. https://doi.org/10.1111/j.1471- 8286.2007.01678.x
Ratti E. (1991) – Catalogo dei coleotteri della laguna di Venezia VI. Lucanidae, Trogidae, Aphodiidae, Scarabaeidae, Melolonthidae, Rutelidae, Dynastidae, Cetoniidae. Lavori Soc. Ven. Scien. Nat., 16: 91-125. Venezia.
Razowski J. (2003) – Tortricidae of Europe vol. 2, Olethreutinae. Bratislava, 301 pp.
Roesler R. U. (1967) – Phycitinien – Studien (Lepidoptera) II. Ent. Z. 77: 273-282.
Robinson S. G. (1976) – The preparation of slides of lepidoptera genitalia with special reference to microlepidoptera. Reprinted from Entomologist’s Gazzette 27: 127-132.
Simonella I. (2006) – Atlante degli ambiti di interesse naturalistico della provincia di Venezia. Provincia di Venezia, Assessorato alle politiche ambientali. Cicero Editore, Venezia: pp. 65-67.
Studiger O. (1879) – Lepidopterenfauna Kleinasien. Horae Soc. ent. ross.:16-195.
Sutter R. (2007) – Neue Arten der Gattung Oegoconia (Autostichidae). Nota lepidopterologica 30 (1): 189-201.
Timossi G. (2009) – I Macrolepidotteri Eteroceri del Parco Nazionale Dolomiti Bellunesi (Lepidoptera Heterocea: Bombyces et Sphinges, Sesiidae, Geometridae, Noctuidae). Frammenti. Conoscere e tutelare la natura bellunese: N.1: 71-93.
Timossi G. (2013) – Contributo alla conoscenza dei Microlepidotteri del Parco Nazionale Dolomiti Bellunesi. Frammenti, conoscere e tutelare la natura bellunese, n° 5: pp. 31-52.
Timossi G. (2014) – Primo contributo alla conoscenza dei Lepidotteri delle prealpi venete. Il Monte Grappa: la valle di Schievenin (Quero, Belluno). Microlepidoptera, bombyces et sphinge. First contribution to the knowledge of lepidoptera of Veneto’s foothill. Massif of Monte Grappa: the Valley of Schievenin (Quero, Belluno). Microlepidoptera, bombyces et sphinge. De Rerum Natura, Quaderni del Museo di Storia Naturale e Archeologia di Montebelluna, n° 7 (2013): pp 5-68.
Trematerra P. (2003) – Catalogo dei Lepidoptera Tortricidae della fauna italiana: geonemia, distribuzione in Italia, note biologiche, identificazione. Bollettino di Zoologia Agraria e Bachicoltura, Ser. II. 35 (suppl. I): 1-270.
Triberti P., Longoturri G., Adami R. & Zanetti A. (2017) – I Lepidotteri della Palude del Busatello” (Verona, Mantova). Memorie Società Entomologica Italiana, 94: 91-135.
Triberti P., Adami R., dall’O M. (2018) – I Lepidottteri. In: Latella L. (ed). La Val Gallina e il Progno Borago. Memorie del Museo Civico di Storia Naturale di Verona, 2. serie. Monografie Naturalistiche 5: 63-70.
Varenne, T. & J. Nel (2013) – Cydia molybdana (Constant, 1884), bona species, stat. restaur. et description de Cydia rymarczyki sp. nov. du sud de la France (Lep. Tortricidae, Olethreutinae). Oreina 21: 9-11.
Zangheri S. (1958) – Osservazioni su alcune interessanti specie di Lepidotteri della laguna di Venezia. Boll. Mus. Civ. St. Nat. di Venezia, 11:19-27.
Sitografia
http://www.boldsystems.org/
http://vnr.unipg.it/habitat/index.jsp
Nuss, M., B. Landry, R. Mally, F. Vegliante, A. Tränkner, F. Bauer, J. Hayden, A. Segerer, R. Schouten, H. Li, T. Trofimova, M. A. Solis, J. De Prins & W. Speidel 2003-2019: Global Information System on Pyraloidea. – www.pyraloidea.org
Pesi (2019) – Pan-European Species directories Infrastructure. Accessed through www.eu-nomen.eu/portal, at 2019-11-27
Stoch F. (2003) – Checklist of the species of italian fauna, on-line version 2.0. Ministero dell’Ambiente e della tutela del territorio e del mare.
