Riassunto
Nella primavera 2015 sono stati effettuati 10 punti di ascolto, ripetuti due volte, per caratterizza-re la comunità ornitica presente sulle praterie sommitali del Monte Cesen (Valdobbiadene, Tv) e cime vicine, poste ad una quota di circa 1.400 m s.l.m. Sono state contattate 17 specie, ciascuna mediamente presente con 5,3 ind. (D.S.= + 5,4). Le specie dominanti (>5%) del totale) sono risultate prispolone Anthus trivialis, allodola Alauda arvensis, cornacchia grigia Corvus cornix, rondone comune Apus apus, culbianco Oenanthe oenanthe; le più diffuse prispolone, allodola e cornacchia grigia. La diversità (H’) è risultata pari a 2,5 e l’equiripartizione (J) a 0,87. Le dif-ferenze tra i valori medi per punto di ascolto di ricchezza (S), abbondanza (N) e diversità (H’) non sono risultate significativamente diverse tra le due sessioni di rilievo. Le densità di territori ottenute sono per allodola e prispolone di 4,1/10 ha, per il culbianco di 1,3 /10 ha e di 0,9/10 ha per l’averla piccola. Le praterie studiate evidenziano un elevato valore per la conservazione di queste e di altre specie tipiche delle praterie sommitali.
Abstract
Breeding bird community in the mountain meadows of the Mon-te Cesen (Veneto Prealps, NE Italy)
In May and June 2015 the bird community of the mountain meadows of the Monte Cesen (1,540 m above sea level; Veneto Prealps), was investigated with the point count method. Seventeen spe-cies were recorded overall; among these, Tree Pipit Anthus trivialis, Skylark Alauda arvensis and Hooded Crow Corvus cornix were the most abundant and the most widespread ones. Diversity (H’) index was 2.5, and evenness (J) 0.87; no significant differences were observed between the two field surveys. The estimated densities for the most abundant grassland species were 4.1 territories/10 ha for Skylark and Tree Pipit, 1.4/10 ha for Weather Oenanthe oenanthe and 0.9/10 ha for Red-backed Shrike Lanius collurio. Mountain prairies in the study area appear to be a stronghold for species, such as Skylark, which are heavily threatened in the lowland farmlands of the Veneto region.
Premessa
Con il termine di praterie sommitali si intendono le formazioni prevalentemen-te erbacee poste alle pendici di complessi collinari o montuosi; in base alla loro genesi vengono suddivise in praterie primarie, la cui presenza è dovuta a fattori naturali edafici e climatici, e praterie secondarie, originatesi per taglio di pre-esistenti formazioni arboree e successivo utilizzo antropico, rappresentato in particolare dallo sfalcio e dal pascolo di ungulati domestici (Buffa & Lasen, 2010). Una recente indagine (Scotton & Cossalter, 2014) indica un’estensione di circa 57.000 ettari per le formazioni erbacee e a prato permanente presenti in ambito collinare e montano dell’intero Veneto.
Anche nel Veneto, come altrove, questi ambienti rivestono particolare interesse naturalistico per la presenza di elementi floristici e faunistici di pregio (Ziliot-to et al., 2004; Buffa & Lasen, 2010). Tuttavia le praterie sommitali sono spes-so in via di degrado o contrazione sia in Italia che nel resto d’Europa, per cause naturali quali la progressiva avanzata del bosco a seguito del cessato o ridotto utilizzo zootecnico o per altre cause di natura antropica, come la frammenta-zione delle superfici conseguente allo sviluppo di reti stradali o di infrastrutture dedicate al tempo libero (Laiolo et al., 2010; Bazzi et al., 2014).
Per quanto concerne l’avifauna, per molte specie di elevato valore conserva-zionistico questi ambienti rappresentano habitat d’elezione sia per la ricerca del nutrimento che per la nidificazione. Nonostante la diffusione ed estensione complessiva che le praterie sommitali hanno nel Veneto, gli studi quali-quanti-tativi sulla loro comunità ornitica sono piuttosto rari; tra i pochi pubblicati, si citano i lavori di Cassol & Dal Farra (2001) e Cassol et al. (2003). I risultati di indagini di campo compiute nel maggio-giugno 2006 nelle praterie sommi-tali del complesso del Monte Zogo-Monte Miliana (Comune di Segusino, TV) sono invece rimasti inediti (Baldin & Scarton, oss. pers.). Numerosi dati or-nitologici riferiti a praterie sommitali e pascoli prealpini si possono trovare in Mezzavilla & Bettiol (2007), mentre sintesi delle conoscenze disponibili sono riportate in alcuni dei numerosi Piani di Gestione elaborati negli anni scorsi per le Zone di Protezione Speciali (ZPS) del Veneto.
La dorsale collinare Cesen-Visentin, estesa per circa 30 km tra il corso del Piave a Ovest e la Valle Lapisina a Est, presenta ampie estensioni di praterie som-mitali, prevalentemente soggette a pascolo bovino od ovi-caprino più o meno intenso; il settore occidentale della dorsale rientra in parte nel SIC IT 3240003 Monte Cesen, mentre la più vasta ZPS IT 3240024 Dorsale prealpina tra Val-dobbiadene e Serravalle (11.622 ha) si estende su buona parte dell’intero setto-re prealpino considerato. Nonostante l’elevato valore naturalistico riconosciuto per questi due Siti Natura 2000, le pubblicazioni scientifiche a carattere orni-tologico che li interessino anche solo in parte sono estremamente scarse; tra le poche si citano Basso et al. (1999), Cassol & Dal Farra (2001) e Mezzavilla et al. (2015).
Con questa nota si intende analizzare la comunità ornitica presente in periodo riproduttivo in un complesso di praterie sommitali circostanti il Monte Cesen (Tv), valutando nel contempo alcuni aspetti metodologici relativi all’applicazio-ne della metodica dei punti di ascolto in tali contesti.
Area di studio e metodi
L’area di studio interessa parte delle estese praterie sommitali che si trovano tra il Monte Cesen (m 1.570 s.l.m.: 45°56’30’’ N, 12°01’28’’ E), il Col Toront (m 1.433) e il Monte Barbaria (1.464 m), nel Comune di Valdobbiadene (Tv) . Si tratta di praterie soggette a pascolo bovino, che si sviluppano sui versanti di questi monti di modesta elevazione e nelle aree meno acclivi tra di loro inter-poste. L’area è caratterizzata da clima oceanico, con inverni freddi, estati tem-perato-calde e piovosità annuale di circa 1.500 mm (Aa.Vv., s.d.). La vegeta-zione dell’area presenta soprattutto componenti acidofile (nardeti, inquadrabili nell’habitat comunitario “*6230 Formazioni erbose a Nardus, ricche di specie, su substrato siliceo delle zone montane e delle zone submontane dell’Europa continentale”) e localmente termofile (seslerio-brometi: “*6210 Formazioni er-bose secche seminaturali e facies coperte da cespugli su substrato calcareo”) come descritto da Buffa & Lasen (2010). Le praterie sono delimitate da boschi di faggio, orno-ostrieti e alcuni rimboschimenti di conifere operati nei decenni scorsi. Piccoli nuclei arbustivi costituiti soprattutto da ginepro Juniperus com-munis e Rosa sp. sono localmente presenti. Sono assenti i corsi d’acqua, men-tre sono presenti diverse pozze d’alpeggio; nell’area insistono alcune malghe (Malga Mariech, Malga Barbaria, Malga Fossazza) e viene praticato il pascolo bovino nei mesi di giugno-settembre. Piuttosto estesa è la rete di sentieri, fre-quentati durante la stagione estiva dagli escursionisti; alcune strade sterrate si dipartono da quella asfaltata che sale dalla località Pianezze verso Malga Ma-riech. I pochi manufatti presenti consistono in malghe, ruderi di stalle e alcuni tralicci per la radiofonia.
Per l’effettuazione dell’indagine è stata utilizzata la metodica dei punti di ascolto, comunemente adottata in indagini ornitologiche quantitative (Gregory et al., 2004). Dopo un sopralluogo preliminare sono stati pertanto ubicati conGarmin GPSmap 76cx dieci punti di ascolto; in ciascuno di essi è stata rilevata l’avifauna presente mediante conteggio di tutti gli individui uditi o osservati entro i 100 m dall’osservatore, per una durata di 10 minuti. Un telemetro Leica Rangemaster LAF 900 è stato utilizzato per delimitare con precisione l’area di rilevamento attorno all’osservatore; pertanto la superficie totale indagata equi-vale a 10 * 3,14 ha = 31,4 ha. Ogni punto era distante, con una sola eccezio-ne, almeno 400 metri da quello più vicino; l’intervallo altitudinale è risultato compreso tra 1.218 e 1.499 m s.l.m. La vegetazione entro i 100 m dai punti di ascolto consisteva in formazioni erbacee per almeno il 90% dell’area, con la rimanente frazione costituita da nuclei arbustivi (ginepro Juniperus communis, rosa canina Rosa canina), singoli esemplari arborei (larice Larix decidua, faggio Fagus sylvatica) e superfici nude con modesti affioramenti rocciosi.
I rilievi ornitologici sono stati effettuati tra le ore 6.30 e le 10.30, con condizioni meteorologiche buone; ciascun punto di ascolto è stato rilevato due volte, il 29 maggio ed il 27 giugno 2015.
I dati ornitologici utilizzati sono bruti, quindi non sono stati in alcun modo ela-borati in funzione della diversa contattabilità delle varie specie, come sarebbe stato possibile utilizzando opportune metodiche (ad es. il distance sampling: Thomas et al., 2010). L’ottima visibilità nell’area di indagine lascia ritenere che la contattabilità sia stata elevata per tutte le specie considerate. Per le specie osservate la nidificazione è stata considerata possibile (individui in canto), pro-babile (coppia; adulti allarmanti) o certa (imbeccata; giovani da poco involati) utilizzando alcune delle categorie proprie dei Progetti Atlante dei Nidificanti (si veda ad esempio Mezzavilla & Bettiol, 2007).
I dati di densità espressi come ind./10 ha presentati per alcune specie tipiche delle praterie, sono stati elaborati scegliendo tra i due valori disponibili quello maggiore, per ovviare almeno in parte alla tendenza del metodo qui utilizzato a sottostimare il numero di individui (Sozio & Battisti, 2009). Poiché si sono considerati solo gli individui in canto o, nel caso di coppie, solo uno dei due adulti, le densità corrispondono a quelle dei territori delle diverse specie.
Per descrivere la comunità ornitica sono stati utilizzati i seguenti parametri: ric-chezza (numero di specie = S), abbondanza (numero di individui = N), indice di diversità di Shannon-Weaver (H’) e indice di equiripartizione J (H’/H’ max, dove H’ max = lnS) (Magurran, 2004). Per ciascuno dei due rilievi sono stati inoltre calcolati per ogni punto di ascolto i valori medi di abbondanza (N med), ricchezza (S med) e diversità (H’ med). Le specie dominanti sono considera-te quelle con frequenza percentuale superiore al 5% del totale dei due rilievi (Turc̄ek, 1957).
La similarità tra i popolamenti ornitologici dei due rilievi è stata valutata con l’indice di Sørensen classico, quindi qualitativo, e con l’indice di Bray-Curtis che corrisponde ad un indice di Sørensen quantitativo (Magurran, 2004). Per verificare se le differenze fra i due rilievi fossero significative si è utilizzato il test ANOSIM (acronimo per ANalysis Of SIMilarities). Il valore, denominato R, che viene generato può formalmente variare tra – 1 e 1, ma in ambito ecologico generalmente assume valori tra 0 e +1 (Scardi, 2009). Un valore di R < 0,25 indica una sostanziale uniformità tra i due insiemi considerati, mentre se R > 0,75 vi è una buona separazione (Liordos, 2010).
L’efficacia del campionamento, intesa come percentuale di specie osservate (Sobs) rispetto al numero di specie statisticamente attese (Sest) è stata valutata utilizzando lo stimatore non parametrico di ricchezza specifica Chao1 (Magur-ran, 2004); questo indice viene di seguito presentato con media e relativo inter-vallo di confidenza asimmetrico.
L’ordinamento tra i vari punti di ascolto è stato effettuato mediante non-Metric Multidimensional Scaling (n-MDS). L’n-MDS consente una rappresentazione grafica delle distanze/dissimilarità tra gli oggetti in pochi assi che riassumono le variabili iniziali; il test produce un indice di stress (R), tanto più piccolo quanto migliore è la rappresentazione ottenuta. Si ritiene in letteratura che valori di R <0,15 rappresentino una buona situazione, mentre valori di R > 0,3 indicano un ordinamento del tutto casuale. Tutte le analisi citate, nonché le curve di accumulazione per specie/punti di ascolto, sono state effettuate mediante i software EstimateS v. 9,1 (Colwell, 2013) e PAST v. 2.9 (Hammer et al., 2001). L’esten-sione delle praterie sommitali nei due Siti Natura 2000 è stata valutata tramite riprese satellitari Google Earth e verificata con visite in campo compiute nella primavera 2015.
Risultati
Nella Tab. 1 si riportano le 17 specie contattate in un raggio di 100 m, considerando tutti i risultati ottenuti nei due rilievi (N=90 individui). Ogni specie è risultata mediamente presente con 5,3 ind. (D.S.= + 5,4), con mediana pari a 4. Le specie dominanti sono risultate cinque: prispolone Anthus trivialis, allodola Alauda arvensis, cornacchia grigia Corvus cornix, rondone comune Apus apus,culbianco Oenanthe oenanthe. Per le prime due e per la quinta è stata accertata la nidificazione durante le indagini di campo. Le specie più diffuse sono state invece prispolone (16 punti di ascolto su 20 complessivamente effettuati, pari all’80%), allodola (75%) e cornacchia grigia (30%). L’analisi dei parametri di comunità indica una diversità (H’) pari a 2,5 ed un’equiripartizione (J) di 0,87.
| Poiana | Buteo buteo | 4 | ,4 |
| Gheppio | Falco tinnunculus | 3 | ,3 |
| Lodolaio | Falco subbuteo | 1 | ,1 |
| Rondone comune | Apus apus | 6 | ,7 |
| Allodola | Alauda arvensis ** |
| 20 |
| Prispolone | Anthus trivialis ** | 22 | ,2 |
| Ballerina bianca | Motacilla alba ** | 4 | ,4 |
| Codirosso spazzacamino | Phoenicurus ochruros ** | 3 | ,3 |
| Stiaccino | Saxicola rubetra* | 3 | ,3 |
| Saltimpalo | Saxicola torquatus* | 4 | ,4 |
| Culbianco | Oenanthe oenanthe ** | 5 | ,6 |
| Cinciallegra | Parus major | 2 | ,2 |
| Averla piccola | Lanius collurio ** | 4 | ,4 |
| Cornacchia grigia | Corvus cornix | 7 | ,8 |
| Corvo imperiale | Corvus corax | 2 | ,2 |
| Fanello | Carduelis cannabina | 2 | ,2 |
| Zigolo giallo | Emberiza citrinella* | 2 | ,2 |
Tab. 1. Frequenze percentuali delle specie contattate entro 100 m dal punto di ascolto (entrambi i rilievi considerati, N=90 indd). Sono state inoltre evidenziate le specie per le quali la nidificazione è possibile (*) o probabile/certa (**) nelle stesse aree.
Tra le specie contattate, l’averla piccola Lanius collurio risulta inclusa nell’allegato 1 della Direttiva CE 147/09 Uccelli, mentre tre (allodola, prispolone e sal-timpalo Saxicola torquatus) sono ritenute “minacciate” nella recente Lista Rossa degli uccelli italiani (Peronace et al., 2012). Una specie, il fanello Carduelis cannabina, risulta avere lo status di SPEC 2, mentre tre (allodola, averla piccola e gheppio Falco tinnunculus) risultano invece SPEC3 secondo la classificazione utilizzata da Birdlife International (2004).
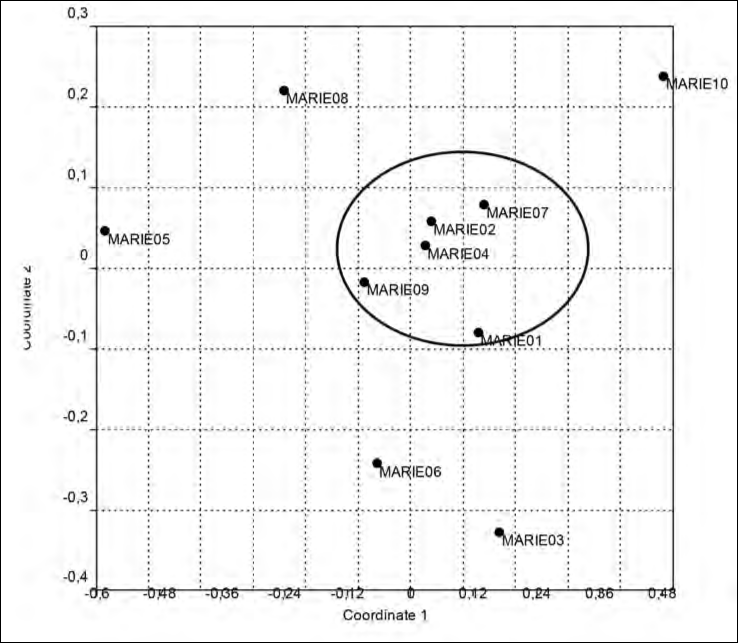
L’individuazione effettuata con n-MDS di gruppi di punti di ascolto tra loro simili è presentata in Fig. 1. Nonostante l’elevata omogeneità ambientale nelle aree campionate, è possibile comunque osservare una discreta differenziazione tra di essi, con un gruppo di cinque punti di ascolto molto vicini tra di loro e che risultano nettamente separati dagli altri. In tre di questi ultimi si concentra la presenza di specie risultate rare nell’indagine, come cinciallegra o rondone comune, elemento che può contribuire a spiegare la suddivisione osservata.
I dati dei due rilievi, considerati separatamente, si prestano ad una serie di ulte-riori considerazioni. La ricchezza specifica è risultata pari a 13 specie nel primo rilievo e a 16 nel secondo; 12 sono le specie contattate in entrambi i rilievi, tra cui allodola, averla piccola, codirosso spazzacamino Phoenicuros ochruros, culbianco e prispolone. Zigolo giallo Emberiza citrinella e stiaccino Saxicola ru-betra sono stati invece osservati solo in uno dei due rilievi. La somiglianza trale comunità ornitiche campionate nei due rilievi, espressa tramite l’indice di Sørensen, risulta essere molto elevata (0,82), mentre l’indice di Bray-Curtis è risultato pari a 0,66. L’analisi ANOSIM restituisce un valore di R=0,003 (con P=0,41) che conferma l’elevata omogeneità tra i popolamenti delle due sessioni di rilievo.
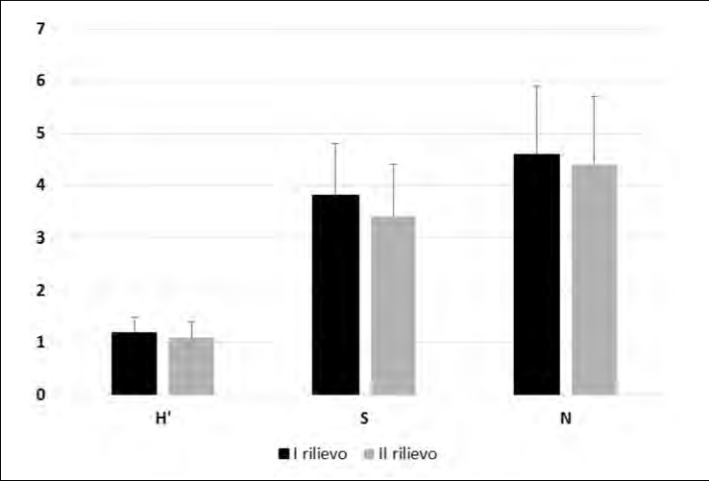
Le differenze tra i valori medi per punto di ascolto di ricchezza (S), abbondanza e diversità (H’) non sono risultate significativamente diverse tra i due rilievi (test t di Student, P>0,05 in tutti i casi: Fig. 2). L’indice Chao1 è risultato pari a 15 specie (I.C.=13-25) per il primo rilievo e 18 specie (I.C.= 16-29) per il se-condo; la proporzione Sobs/Sest varia quindi tra l’80% del primo rilievo e l’83% del secondo.
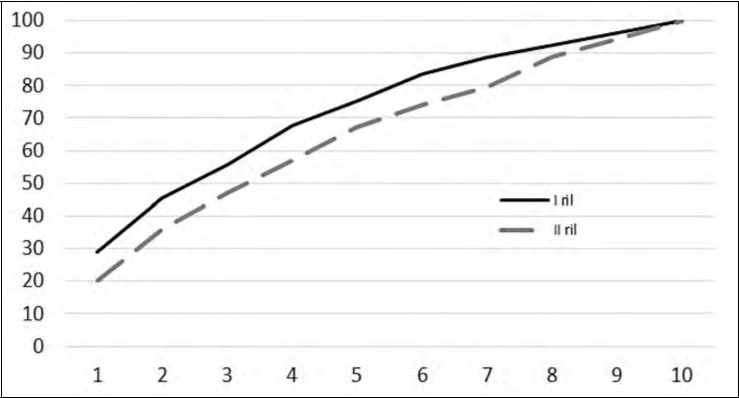
Le due curve di accumulazione specie-punto di ascolto sono simili tra i due rilievi ed evidenziano che già con otto punti di ascolto si raggiunge circa il 90% di tutte le specie complessivamente contattate in un campionamento.
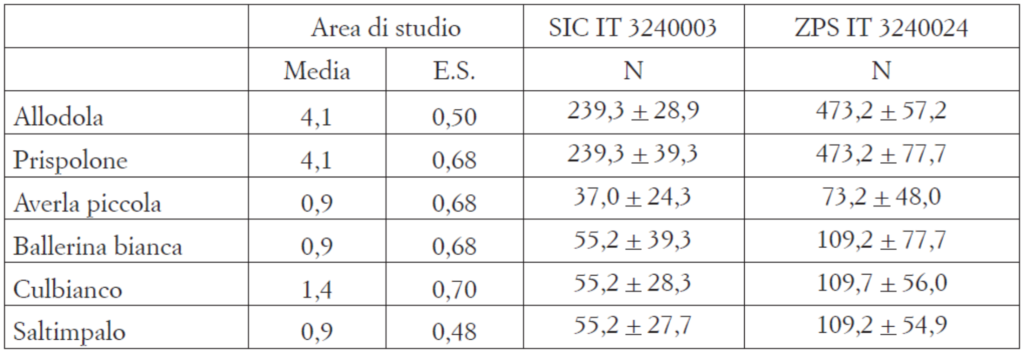
Le densità dei territori per le specie presenti in almeno tre punti di ascolto è riportata in Tab. 2; i valori nettamente più elevati si riferiscono ad allodola e prispolone (4,1/10 ha), seguiti dal culbianco (1,3/10 ha). Poiché è stata stimata la presenza di 578 ettari di praterie sommitali per il SIC Monte Cesen e di 1.143 ha per l’intera ZPS Valdobbiadene-Serravalle, la tab. 2 riporta il numero di ter-ritori delle sei specie per ciascuno dei due Siti, tramite semplice estrapolazione delle densità ottenute nell’area di studio.
Conclusioni
Nell’area di indagine la comunità ornitica è risultata composta da 17 specie, valore modesto in senso assoluto ma comparabile a quelli rilevati in ambienti si-mili italiani. Ad esempio, Laiolo et al. (2004) censiscono 18 specie in aree inter-ne al Parco Nazionale del Gran Paradiso, mentre Casale & Brambilla (2010) ne segnalano 29 in aree di studio situate nel Parco Nazionale Val Grande. Sia Lorenzetti et al. (2004) che Plini (1996) citano invece 22 specie per praterie sommitali del Lazio, mentre 28 specie furono rilevate da Perna & Gaibani (2008) in praterie dell’Appennino marchigiano. In praterie sommitali sul Mon-te Miliana, Baldin & Scarton (oss. pers.) osservarono 22 specie; Cassol & Dal Farra (2001) citano la presenza di 26 specie nei pascoli del Monte Faverghera (Bl), mentre Cassol et al. (2003) elencano 39 specie per ambienti sommitali del Parco Nazionale Dolomiti Bellunesi. Quest’ultimo valore, piuttosto elevato se confrontato con gli altri, si spiega probabilmente con la presenza oltre alle praterie di altre tipologie ambientali, quali mughete, alnete e rocce nude, nelle aree di studio.
Le specie nidificanti (possibili, probabili e certe) nei dintorni dei punti di cam-pionamento sono risultate nove; simili i valori osservati da Cassol & Dal Farra (2001) e da Baldin & Scarton (oss. pers). Escludendo le specie che certamente non nidificano nelle praterie sommitali (cornacchia grigia e rondone comune), la comunità ornitica delle praterie del Monte Cesen risulta dominata da prispo-lone, allodola e culbianco; le buone densità rilevate per l’allodola confermano come gli ambienti di prateria sommitale costituiscano uno dei pochi habitat an-cora idonei per questa specie, che nel Veneto è in forte rarefazione soprattutto negli agroecosistemi di pianura (Bon et al., 2013; Scarton, 2016).
I risultati indicano come lo stiaccino, anch’essa specie caratteristica delle prate-rie sommitali, sia risultato invece poco diffuso e poco abbondante nell’area di indagine; anche nelle praterie sommitali del Monte Miliana nel maggio-giugno 2006 (Baldin & Scarton, oss. pers.) la specie non era stata rilevata, a conferma della sua rarità nel settore prealpino trevigiano, come riportato da Mezzavilla & Bettiol (2007). L’apparente assenza dello spioncello sia dalle praterie qui considerate che da quelle investigate in passato sul Monte Miliana contrasta con l’accertata nidificazione riportata per le pendici del Monte Cesen da Mez-zavilla & Bettiol (2007); è probabile che ciò sia dovuto ad una densità molto bassa della specie o a sue forti fluttuazioni interannuali. L’assenza del sordone dall’area di studio è invece un’ulteriore conferma della rarità di questa specie come nidificante nell’intera provincia di Treviso (Mezzavilla & Bettiol, 2007). Interessanti le osservazioni, seppure limitate a pochi punti, dell’averla piccola; questa specie nelle praterie del Monte Cesen è stata rilevata in macchie arbu-stive, sempre di modeste dimensioni, dominate da rosa canina e spin cervino Rhamnus cathartica; ciò conferma l’importanza della presenza, o del manteni-mento, di questi nuclei arbustivi in aree soggette per la maggior parte a pascolo. Le praterie sommitali campionate nella presente indagine si confermano aree di caccia per alcuni rapaci diurni quali gheppio, poiana Buteo buteo e il più raro lodolaio Falco subbuteo; nel giugno 2015 al difuori delle osservazioni standar-dizzate è stato osservato in un’occasione anche il biancone Circaetus gallicus, specie nidificante molto rara nell’intero arco delle Prealpi venete (Bon et al. 2013).
I valori di densità delle specie più diffuse ottenuti per le praterie sommitali del Monte Cesen sono circa il doppio di quanto osservato da Cassol & Dal Farra (2001) con il metodo del mappaggio per allodola e prispolone, mentre sono poco più bassi per il culbianco. In aree prealpine della Lombardia, Bazzi et al. (2014) rilevarono valori confrontabili con quelli della presente indagine sia per le tre specie citate che per l’averla piccola. Le stime relative al numero di terri-tori ottenute per ciascuno dei due Siti Natura 2000 delle Prealpi trevigiane, an-corché preliminari e meritevoli di approfondimento, indicano l’importanza che questi comprensori hanno per la conservazione di specie tipiche delle praterie. L’utilizzo dei punti di ascolto è risultato efficace per campionare la comunità ornitica dell’ambiente in esame; anche in ragione della modesta ricchezza spe-cifica e della non elevata diversità, si può suggerire una densità efficace di un punto di ascolto ogni 50 ha circa per indagini in aree simili. Il rilievo effettuato in giugno ha evidenziato più specie di quello effettuato in maggio, con un nu-mero confrontabile di individui; nel complesso vi è stata alta somiglianza tra i due popolamenti. Per indagini speditive finalizzate alla caratterizzazione della comunità ornitica un solo rilievo nella seconda metà di giugno appare pertan-to sufficiente, purché le condizioni meteo siano ottimali per l’esecuzione delle indagini di campo.
Anche nel Veneto, come nel resto d’Italia, dal 2000 è attivo il progetto MITO2000 (Monitoraggio ITaliano Ornitologico), un programma di monitoraggio dell’avi-fauna nidificante che ha il principale obiettivo di fornire indicazioni sugli anda-menti di popolazione nel tempo e di calcolare indicatori aggregati (Fornasari et al., 2010). Tra questi indicatori, dal 2010 viene calcolato a livello nazionalequello relativo agli uccelli delle praterie montane, in considerazione della speci-ficità della comunità ornitica che le utilizza per nidificare e della loro prevedibi-le sensibilità ai cambiamenti climatici già in atto; i più recenti risultati relativi al 2000-2014 indicano un trend fortemente negativo per l’insieme di queste specie (Rete Rurale & Lipu, 2015).
L’elaborazione di un indicatore per l’avifauna delle praterie montane non è attualmente possibile per il Veneto, dato l’esiguo numero di punti di ascolto disponibili per questi ambienti; ciò impedisce di avere informazioni affidabili sugli andamenti delle specie che li utilizza nel periodo riproduttivo. L’ubicazione di una rete di nuovi punti consentirebbe pertanto, con modesto sforzo logistico, di acquisire dati di fondamentale importanza sul medio-lungo periodo, anche per valutare i possibili effetti delle trasformazioni ambientali in atto in territori di elevato valore ornitologico e spesso inclusi in Siti della rete Natura 2000.
Ringraziamenti
L’Associazione Faunisti Veneti ha messo a disposizione un telemetro laser; l’amico Francesco Mezzavilla ha cortesemente rivisto il testo.
Bibliografia
Aa.Vv. (s.d.) – Piano di gestione ZPS IT32240024 Dorsale prealpina tra Valdobbiadene e Serravalle. Internet: www.prealpitrevigiane.it
Basso E., Martignago G., Silveri G., Mezzavilla F. (1999) – Censimenti del Re di Quaglie Crex crex nelle Prealpi Venete Orientali. Anni 1994-1998. Avocetta, 23:115
Bazzi G., Foglini C., Brambilla M., Saino N., Rubolini D. (2014) – Habitat management effects on Prealpine grassland bird communities. Italian Journal of Zoology: 1-11.
Birdlife International (2004) – Birds in the European Union: a status assessment.
BirdLife International, Wageningen,The Netherlands. Bird Life International.
Bon M., Mezzavilla F., Scarton F. (eds.) (2013) – Carta delle Vocazioni Faunistiche del Veneto. Regione del Veneto-Associazione Faunisti Veneti. 586 pp.
Buffa G., Lasen C. (2010) – Atlante dei siti Natura 2000 del Veneto. Regione del Ve-neto. Regione del Veneto – Direzione Pianificazione Territoriale e Parchi, pp. 1-394.
Casale F., Brambilla M. (2010) – L’avifauna nidificante negli ambienti aperti del Parco Nazionale della Val Grande. Ente Parco Nazionale della Val Grande e Fondazione Lombardia per l’Ambiente.
Cassol M., Bertoldin S., D’Alberte G., Dal Farra A. (2003) – Uccelli nidificanti nel Parco Nazionale Dolomiti Bellunesi. La comunità degli ambienti al di sopra del limite del bosco nelle Vette Feltrine e sul Monte Cirvoi. In Bon M., Fracasso G., Dal Lago A. (red). Atti 4° Convegno Faunisti Veneti. Natura Vicentina, 7: 159-162.
Cassol M., Dal Farra A. (2001) – Studio di una comunità di Passeriformi su di un’area a pascolo delle Prealpi Bellunesi (M. Faverghera – Bl). In Bon M. e Scarton F. Atti III Convegno Faunisti Veneti: 210-213.
Colwell R.K. (2013) – EstimateS. Statistical estimation of species richness and shared species from samples. Version 9. URL: purl.oclc.org/estimates.
Fornasari L., Londi G., Buvoli G., Tellini Florenzano G., La Gioia G., Pedrini
P., Brichetti P., de Carli E. (eds.) (2010) – Distribuzione geografica e ambientale degli uccelli comuni nidificanti in Italia, 2000-2004 (dati del progetto MITO2000). Avocetta, 34: 1-224.
Gregory R.D., Gibbons D.W., Donald P.F. (2004) – Bird census and survey techniques. In: Sutherland W.J., Newton I., Green R.E. [eds.]. Bird Ecology and Conservation; a Handbook of Techniques. Oxford University Press, Oxford: 17-56.
Hammer Ø., Harper D.A.T., Ryan P.D. (2001) – PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica, 4: 9.
Web: http://folk.uio.no/ohammer/past.
Laiolo P., Dondero F., Ciliento E., Rolando A. (2004) – Consequences of pastoral abandonment for the structure and diversity of the alpine avifauna. Journal of Ap-plied Ecology,41:294-304.
Liordos V. (2010) – Foraging Guilds of Waterbirds Wintering in a Mediterranean Coastal Wetland. Zoological Studies, 49: 311-323.
Lorenzetti E., Ukmar E., Battisti C. (2004) – Comunità ornitiche nidificanti nel sito di importanza comunitaria “Monte Guadagnolo” (Monti Prenestini, Italia centrale). Alula,11: 105-112.
Magurran A. (2004) – Measuring biological diversity. Blackwell Publishing, Malden. Mezzavilla F., Martignago G., Silveri G. (2015) – Monitoraggio del re di quaglie Crex crex nelle Prealpi venete orientali. In Pedrini P., Rossi F., Bogliani G., SerraL. & Sustersic A. (a cura di) 2015. XVII Convegno Italiano di Ornitologia: Atti del convegno di Trento. Ed. MUSE, 176 pp.
Mezzavilla F. , Bettiol K. (2007) – Nuovo Atlante degli Uccelli nidificanti in provincia di Treviso. (2003-2006). Associazione Faunisti Veneti. Pp. 198.
Perna P., Gaibani G. (2008) – Studio sull’ecologia dell’ornitofauna nell’Appennino umbro-marchigiano. In Roscelli F. (ed.). Atti del Convegno Uccelli di prati e pascoli: stato e prospettive di conservazione. Natura Modenese 8: 3-34.
Peronace V., Cecere J., Gustin M., Rondinini C. (2012) – Lista Rossa 2011 degli Uccelli Nidificanti in Italia. Avocetta, 36: 11-58.
Plini P. (1996) – Analisi quantitativa dell’avifauna nidificante in un ambiente ad elevata eterogeneità: la conca di Amatrice e l’altipiano di Campotosto (Parco Nazionale Gran Sasso – Monti della Laga. Alula, 3: 68-77.
Rete Rurale Nazionale & Lipu (2015) – Uccelli comuni in Italia. Aggiornamento degli andamenti di popolazione e del Farmland Bird Index per la Rete Rurale Nazionale dal 2000 al 2014.
Scardi M. (2009) – Tecniche di analisi dei dati in ecologia. Dipartimento di Biologia, Università di Roma. Internet: http://www.mare-net.com/mscardi. Ultimo accesso: 15 dicembre 2014.
Scarton F. (2016) – L’avifauna presente in periodo riproduttivo in un’area ad agricol-tura intensiva del Veneto. Alula, 23(1-2): 99-110.
Scotton M., Cossalter S. (2014) – Praterie seminaturali ricche di specie nella pianu-ra veneta. Distribuzione e valorizzazione negli interventi di inerbimento e restauro ecologico. Veneto Agricoltura.
Sozio G., Battisti C. (2009) – Comunità ornitiche nidificanti in un mosaico ambientale antropizzato (litorale laziale, Italia centrale). Ecologia Urbana, 21: 3-7.
Thomas L., Buckland S. T., Rexstad E. A., Laake J. L., Strindberg S., Hedley S. L., Burnham K. P. (2010) – Distance software: Design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology, 47: 5-14.
Turc̄ek F.J. (1957) – The bird succession in the conifer plantations on mat-grass land in Slovakia (CSR). Ibis, 99: 587-593
Ziliotto U. (coord.), Andrich O., Lasen C., Ramanzin M. (2004) – Tratti essenziali del-la tipologia veneta dei pascoli di monte e dintorni. Regione del Veneto, Accademia Italiana di Scienze Forestali, (Venezia).
